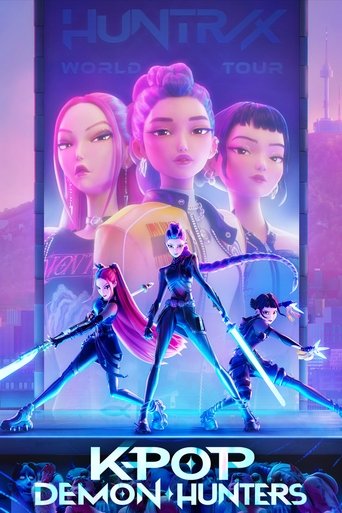







| Dilophosaurus | |
|---|---|

| |
| Rekonstruksi cetakan spesimen holotipe (UCMP 37302) dalam posisi saat terkubur, Royal Ontario Museum | |
| Klasifikasi ilmiah | |
| Kerajaan: | Animalia |
| Filum: | Chordata |
| Klad: | Dinosauria |
| Klad: | Saurischia |
| Klad: | Theropoda |
| Klad: | Neotheropoda |
| Genus: | †Dilophosaurus Welles, 1970 |
| Spesies: | †D. wetherilli
|
| Nama binomial | |
| †Dilophosaurus wetherilli (Welles, 1954)
| |
| Sinonim | |
| |
Dilophosaurus (/daɪˌloʊfəˈsɔːrəs, -foʊ-/[1] dy-LOH-fə-SOR-əs, -foh-) adalah sebuah genus dinosaurus teropoda yang hidup di wilayah yang sekarang menjadi Amerika Utara pada masa Jura Awal, sekitar 186 juta tahun yang lalu. Tiga kerangka ditemukan di Arizona utara pada tahun 1940, dan dua spesimen yang paling terawetkan dikumpulkan pada tahun 1942. Spesimen yang paling lengkap menjadi holotipe untuk spesies baru dalam genus Megalosaurus, yang diberi nama M. wetherilli oleh Samuel P. Welles pada tahun 1954. Welles menemukan kerangka yang lebih besar di wilayah yang sama dan berasal dari spesies yang sama pada tahun 1964. Menyadari bahwa tengkoraknya memiliki jambul, ia memindahkan spesies tersebut ke dalam genus baru Dilophosaurus pada tahun 1970, sebagai Dilophosaurus wetherilli. Nama genus ini berarti "kadal berjambul dua", dan nama spesiesnya didedikasikan untuk menghormati John Wetherill, seorang penjelajah dan arkeolog amatir. Spesimen-spesimen lain telah ditemukan sejak saat itu, termasuk spesimen bayi. Jejak kaki fosil juga telah dikaitkan dengan hewan ini, termasuk jejak saat beristirahat. Spesies lain, Dilophosaurus sinensis dari Tiongkok, diberi nama pada tahun 1993, namun kemudian diketahui merupakan bagian dari genus Sinosaurus.
Dengan panjang sekitar 7 m (23 ft) dan berat sekitar 400 kg (880 pon), Dilophosaurus adalah salah satu dinosaurus predator besar terawal dan hewan darat terbesar yang diketahui di Amerika Utara pada masa itu. Tubuhnya ramping dan berkerangka ringan, serta tengkoraknya proporsional besar, namun rapuh. Moncongnya sempit, dan rahang atasnya memiliki celah atau lekukan di bawah lubang hidung. Hewan ini memiliki sepasang jambul melengkung yang memanjang di tengkoraknya; bentuk utuhnya tidak diketahui, tetapi kemungkinan diperbesar oleh keratin. Tulang rahang bawahnya ramping dan tipis di bagian depan, namun menebal di bagian belakang. Giginya panjang, melengkung, tipis, dan memipih di bagian samping. Gigi di rahang bawah jauh lebih kecil daripada gigi di rahang atas. Sebagian besar giginya memiliki gerigi di tepi depan dan belakangnya. Lehernya panjang, dan tulang belakangnya berongga, serta sangat ringan. Lengannya kuat, dengan tulang lengan atas yang panjang dan ramping. Tangannya memiliki empat jari; jari pertama pendek namun kuat dan memiliki cakar yang besar, dua jari berikutnya lebih panjang dan ramping dengan cakar yang lebih kecil; jari keempat merupakan organ vestigial. Tulang pahanya masif, kakinya kokoh, dan jari-jari kakinya memiliki cakar yang besar.
Dilophosaurus telah dianggap sebagai anggota dari famili Dilophosauridae bersama dengan Dracovenator, sebuah kelompok yang ditempatkan di antara Coelophysidae dan teropoda di masa berikutnya, namun beberapa peneliti belum menemukan dukungan untuk pengelompokan ini. Dilophosaurus diduga merupakan hewan aktif dan bipedal, dan kemungkinan memburu hewan besar; ia juga dapat memangsa hewan-hewan yang lebih kecil dan ikan. Karena rentang pergerakan yang terbatas dan pendeknya tungkai depan, mulutnya kemungkinan menjadi bagian pertama yang bersentuhan dengan mangsa. Fungsi dari jambul tersebut tidak diketahui; struktur tersebut terlalu lemah untuk digunakan bertarung, tetapi mungkin digunakan untuk pertunjukan visual, seperti pengenalan spesies dan seleksi seksual. Hewan ini mungkin tumbuh dengan cepat, mencapai tingkat pertumbuhan 30 hingga 35 kg (66 hingga 77 pon) per tahun pada awal kehidupannya. Spesimen holotipe memiliki banyak paleopatologi, termasuk luka yang telah sembuh dan tanda-tanda anomali perkembangan. Dilophosaurus diketahui dari Formasi Kayenta, dan hidup berdampingan dengan dinosaurus lain seperti Scutellosaurus dan Sarahsaurus. Hewan ini ditetapkan sebagai dinosaurus negara bagian Connecticut berdasarkan jejak-jejak kaki yang ditemukan di sana. Dilophosaurus ditampilkan dalam novel Jurassic Park dan adaptasi filmnya, di mana hewan ini diberi kemampuan fiktif untuk meludahkan bisa dan mengembangkan jumbai leher, serta digambarkan berukuran lebih kecil dari hewan aslinya.
Sejarah penemuan
sunting
Pada musim panas tahun 1942, ahli paleontologi Charles L. Camp memimpin rombongan lapangan dari University of California Museum of Paleontology (UCMP) untuk mencari fosil vertebrata di County Navajo di Arizona utara. Berita mengenai hal ini menyebar di kalangan penduduk asli Amerika di sana, dan seorang Navajo bernama Jesse Williams membawa tiga anggota ekspedisi tersebut ke beberapa tulang fosil yang telah ia temukan pada tahun 1940. Area tersebut merupakan bagian dari Formasi Kayenta, sekitar 32 km (20 mi) di sebelah utara Cameron dekat Tuba City di Reservasi Indian Navajo. Tiga kerangka dinosaurus ditemukan di dalam serpih keunguan, tersusun dalam bentuk segitiga, dengan panjang sekitar 9,1 m (30 ft) di satu sisinya. Kerangka pertama hampir lengkap, hanya kehilangan bagian depan tengkorak, sebagian panggul, dan beberapa tulang belakang. Kerangka kedua sangat tererosi, mencakup bagian depan tengkorak, rahang bawah, beberapa tulang belakang, tulang tungkai, dan tangan yang terartikulasi. Kerangka ketiga sangat tererosi sehingga hanya menyisakan fragmen-fragmen tulang belakang. Kerangka pertama yang berkondisi baik dibungkus dalam balok gips setelah dikerjakan selama 10 hari dan dimuat ke atas truk, kerangka kedua mudah dikumpulkan karena hampir seluruhnya telah melapuk keluar dari tanah, namun kerangka ketiga hampir hancur tak bersisa.[2][3][4]
Spesimen pertama yang hampir lengkap dibersihkan dan dirangkai di UCMP di bawah pengawasan ahli paleontologi Wann Langston, sebuah proses yang memakan waktu dua tahun oleh tiga orang. Kerangka tersebut dipasang di dinding dalam bentuk relief rendah, dengan ekor melengkung ke atas, leher diluruskan, dan kaki kiri diangkat agar lebih terlihat, namun sisa kerangka lainnya dipertahankan dalam posisi seperti saat terkubur. Karena tengkoraknya hancur, bagian ini direkonstruksi berdasarkan bagian belakang tengkorak spesimen pertama dan bagian depan spesimen kedua. Panggulnya direkonstruksi menyerupai panggul Allosaurus, dan kaki-kakinya juga ikut direkonstruksi. Pada saat itu, ini adalah salah satu kerangka dinosaurus teropoda yang paling awet, meskipun tidak lengkap. Pada tahun 1954, ahli paleontologi Samuel P. Welles, yang merupakan bagian dari kelompok yang menggali kerangka-kerangka tersebut, secara awal mendeskripsikan dan menamai dinosaurus ini sebagai spesies baru dalam genus yang sudah ada yaitu Megalosaurus, M. wetherilli. Spesimen yang hampir lengkap (dikatalogkan sebagai UCMP 37302) dijadikan holotipe untuk spesies ini, dan spesimen kedua (UCMP 37303) dijadikan paratipe. Nama spesifiknya didedikasikan untuk menghormati John Wetherill, seorang anggota dewan Navajo yang digambarkan oleh Welles sebagai "penjelajah, sahabat para ilmuwan, dan pedagang tepercaya". Keponakan Wetherill, Milton, adalah orang yang pertama kali memberi tahu pihak ekspedisi mengenai fosil-fosil tersebut. Welles menempatkan spesies baru ini ke dalam Megalosaurus karena proporsi tungkainya yang mirip dengan M. bucklandii, dan karena ia tidak menemukan perbedaan besar di antara keduanya. Pada saat itu, Megalosaurus digunakan sebagai "takson keranjang sampah", di mana banyak spesies teropoda ditempatkan di dalamnya, tanpa memandang umur atau lokasi penemuannya.[2][5][3][6]
Welles kembali ke Tuba City pada tahun 1964 untuk menentukan umur Formasi Kayenta (sebelumnya diusulkan berumur Trias Akhir, sedangkan Welles berpendapat formasinya berumur Jura Awal hingga Jura Tengah), dan menemukan kerangka lain sekitar 400 m (1⁄4 mi) di selatan lokasi penemuan spesimen tahun 1942. Spesimen yang hampir lengkap ini (dikatalogkan sebagai UCMP 77270) dikumpulkan dengan bantuan William J. Breed dari Museum of Northern Arizona dan pihak lainnya. Selama masa preparasi spesimen ini, menjadi jelas bahwa individu ini adalah M. wetherilli yang berukuran lebih besar, dan bahwa hewan ini memiliki dua jambul di bagian atas tengkoraknya. Karena berupa lempengan tulang yang tipis, salah satu jambul pada awalnya dikira sebagai bagian dari sisi kiri tengkorak yang hilang, yang mungkin telah ditarik keluar dari posisinya oleh seekor pemakan bangkai. Ketika struktur tersebut dipastikan sebagai sebuah jambul, disadari pula bahwa terdapat jambul pasangannya di sisi kiri, karena jambul kanan berada di sebelah kanan garis tengah tengkorak, dan berbentuk cekung di sepanjang bagian tengahnya. Penemuan ini mengarah pada pemeriksaan ulang terhadap spesimen holotipe, yang mana ditemukan memiliki dasar dari dua tulang tipis yang memanjang ke atas, yang hancur menjadi satu. Tulang-tulang ini juga merupakan jambul, namun sebelumnya diasumsikan sebagai bagian dari tulang pipi yang salah tempat. Kedua spesimen tahun 1942 tersebut juga diketahui merupakan individu remaja, sedangkan spesimen tahun 1964 merupakan individu dewasa, dengan ukuran sekitar sepertiga lebih besar dari spesimen lainnya.[2][7][8] Welles kemudian mengenang bahwa ia menganggap keberadaan jambul tersebut sama tak terduganya dengan menemukan "sayap pada seekor cacing".[9]
Genus baru dan penemuan selanjutnya
sunting

Welles dan seorang asistennya kemudian mengoreksi rangkaian kerangka dinding spesimen holotipe berdasarkan kerangka yang baru, dengan memulihkan jambulnya, membuat ulang panggulnya, membuat tulang rusuk lehernya lebih panjang, dan menempatkannya lebih berdekatan. Setelah mempelajari kerangka teropoda Amerika Utara dan Eropa, Welles menyadari bahwa dinosaurus ini bukan bagian dari Megalosaurus, dan membutuhkan nama genus baru. Pada saat itu, belum ada teropoda lain dengan jambul memanjang besar di kepalanya yang diketahui, sehingga dinosaurus ini menarik perhatian para ahli paleontologi. Cetakan dari spesimen holotipe pun dibuat, dan salinan fiberglass-nya didistribusikan ke berbagai pameran; untuk memudahkan pelabelan salinan-salinan ini, Welles memutuskan untuk menamai genus baru ini dalam sebuah catatan singkat, alih-alih menunggu hingga publikasi deskripsi yang terperinci. Pada tahun 1970, Welles menciptakan nama genus baru Dilophosaurus, dari kata Yunani di (δι) yang berarti "dua", lophos (λόφος) yang berarti "jambul", dan sauros (σαυρος) yang berarti "kadal": "kadal berjambul dua". Welles menerbitkan deskripsi osteologis yang mendetail mengenai Dilophosaurus pada tahun 1984, namun tidak menyertakan spesimen tahun 1964 karena ia menduga spesimen tersebut berasal dari genus yang berbeda.[2][7][11][8][10] Dilophosaurus merupakan teropoda pertama yang dikenal luas dari masa Jura Awal, dan masih menjadi salah satu contoh paling terawetkan dari zaman tersebut.[5]
Pada tahun 2001, ahli paleontologi Robert J. Gay mengidentifikasi sisa-sisa dari setidaknya tiga spesimen Dilophosaurus baru (jumlah ini didasarkan pada keberadaan tiga fragmen tulang pubis dan dua tulang paha dengan ukuran yang berbeda) dalam koleksi Museum of Northern Arizona. Spesimen-spesimen tersebut ditemukan pada tahun 1978 di Rock Head Quadrangle, sejauh 190 km (120 mi) dari tempat spesimen aslinya ditemukan, dan sebelumnya diberi label sebagai "teropoda besar". Meskipun sebagian besar materialnya rusak, penemuan ini signifikan karena mencakup elemen-elemen yang tidak terawetkan pada spesimen sebelumnya, termasuk sebagian panggul dan beberapa tulang rusuk. Beberapa elemen dalam koleksi tersebut merupakan bagian dari spesimen bayi (MNA P1.3181), contoh termuda yang diketahui dari genus ini, dan salah satu teropoda bayi terawal yang diketahui dari Amerika Utara, yang hanya didahului oleh beberapa spesimen Coelophysis. Spesimen remaja tersebut mencakup sebagian tulang humerus, sebagian tulang betis, dan fragmen gigi.[12] Pada tahun 2005, ahli paleontologi Ronald S. Tykoski mengaitkan sebuah spesimen (TMM 43646-140) dari Gold Spring, Arizona, ke dalam genus Dilophosaurus, namun pada tahun 2012, ahli paleontologi Matthew T. Carrano dan rekan-rekannya menemukan bahwa spesimen tersebut berbeda dalam beberapa detail.[13][14]
Pada tahun 2020, ahli paleontologi Adam D. Marsh dan Timothy B. Rowe mendeskripsikan ulang Dilophosaurus secara komprehensif berdasarkan spesimen yang telah diketahui saat itu, termasuk spesimen UCMP 77270 yang belum pernah dideskripsikan semenjak 1964. Mereka juga mengeluarkan beberapa spesimen yang sebelumnya telah dikaitkan, karena menganggapnya terlalu terfragmentasi untuk diidentifikasi, dan merelokasi lokasi penggalian spesimen tipe dengan bantuan kerabat Jesse Williams.[6][15] Dalam sebuah wawancara, Marsh menyebut Dilophosaurus sebagai "dinosaurus kurang dikenal yang paling terkenal", karena hewan ini kurang dipahami meskipun telah ditemukan 80 tahun sebelumnya. Masalah utamanya adalah penelitian spesimen sebelumnya tidak memperjelas bagian mana yang merupakan fosil asli dan mana yang direkonstruksi menggunakan gips, padahal peneliti selanjutnya hanya mengandalkan monograf Welles tahun 1984 untuk studi lanjutan, sehingga mengaburkan pemahaman mengenai anatomi dinosaurus tersebut. Marsh menghabiskan waktu tujuh tahun untuk mempelajari spesimen guna mengklarifikasi masalah seputar dinosaurus ini, termasuk dua spesimen yang ditemukan dua dekade sebelumnya oleh Rowe, penasihat Ph.D.-nya.[16]
Spesies yang sebelumnya dikaitkan
sunting
Pada tahun 1984, Welles mengusulkan bahwa spesimen tahun 1964 (UCMP 77270) bukan merupakan bagian dari Dilophosaurus, melainkan genus baru, berdasarkan perbedaan pada tengkorak, tulang belakang, dan tulang paha. Ia berpendapat bahwa kedua genus tersebut memiliki jambul, tetapi bentuk pasti dari jambul pada Dilophosaurus tidak diketahui.[2] Welles meninggal pada tahun 1997, sebelum ia sempat menamai dinosaurus yang diduga baru ini, dan gagasan bahwa keduanya merupakan genus yang terpisah umumnya telah diabaikan atau dilupakan sejak saat itu.[5] Pada tahun 1999, ahli paleontologi amatir Stephan Pickering menerbitkan secara pribadi nama baru Dilophosaurus "breedorum" berdasarkan spesimen tahun 1964, dinamai untuk menghormati Breed, yang telah membantu mengumpulkannya. Nama ini dianggap sebagai sebuah nomen nudum, nama terpublikasi yang tidak valid, dan Gay pada tahun 2005 menunjukkan bahwa tidak ada perbedaan signifikan antara D. "breedorum" dan spesimen D. wetherilli lainnya.[17][18] Pada tahun 2012, Carrano dan rekan-rekannya menemukan perbedaan antara spesimen tahun 1964 dan spesimen holotipe, tetapi menganggapnya sebagai variasi antarindividu alih-alih spesies yang berbeda.[13] Ahli paleontologi Christophe Hendrickx dan Octávio Mateus mengusulkan pada tahun 2014 bahwa spesimen-spesimen yang diketahui mungkin merepresentasikan dua spesies Dilophosaurus berdasarkan ciri tengkorak yang berbeda dan pemisahan stratigrafi, sambil menunggu deskripsi menyeluruh mengenai spesimen-spesimen yang dikaitkan.[19] Marsh dan Rowe menyimpulkan pada tahun 2020 bahwa hanya terdapat satu takson di antara spesimen-spesimen Dilophosaurus yang diketahui, dan bahwa perbedaan di antara mereka disebabkan oleh tingkat kematangan dan pengawetan yang berbeda. Mereka juga tidak menemukan pemisahan stratigrafi yang substansial di antara spesimen-spesimen tersebut.[6]
Kerangka teropoda yang hampir lengkap (KMV 8701) ditemukan di Formasi Lufeng, di Provinsi Yunnan, Tiongkok, pada tahun 1987. Spesimen ini mirip dengan Dilophosaurus, dengan sepasang jambul dan celah yang memisahkan pramaksila dari maksila, tetapi berbeda dalam beberapa detail. Ahli paleontologi Shaojin Hu menamainya sebagai spesies baru Dilophosaurus pada tahun 1993, D. sinensis (dari bahasa Yunani Sinai, merujuk pada Tiongkok).[20] Pada tahun 1998, ahli paleontologi Matthew C. Lamanna dan rekan-rekannya menemukan bahwa D. sinensis identik dengan Sinosaurus triassicus, seekor teropoda dari formasi yang sama yang dinamai pada tahun 1940.[21] Kesimpulan ini dikonfirmasi oleh ahli paleontologi Lida Xing dan rekan-rekannya pada tahun 2013, dan meskipun ahli paleontologi Guo-Fu Wang dan rekan-rekannya sepakat bahwa spesies tersebut merupakan bagian dari Sinosaurus pada tahun 2017, mereka menyarankan bahwa itu mungkin merupakan spesies terpisah, S. sinensis.[22][23]
Deskripsi
sunting
Dilophosaurus adalah salah satu dinosaurus predator besar terawal, seekor teropoda berukuran sedang, meskipun kecil jika dibandingkan dengan beberapa teropoda di masa berikutnya.[2][5] Hewan ini juga merupakan hewan darat terbesar yang diketahui dari Amerika Utara selama masa Jura Awal.[6] Bertubuh ramping dan berkerangka ringan, ukurannya sebanding dengan seekor beruang cokelat.[5][24][25] Spesimen terbesar yang diketahui memiliki berat sekitar 400 kilogram (880 pon), panjang sekitar 7 meter (23 ft), dan panjang tengkoraknya adalah 590 milimeter (23+1⁄4 in). Spesimen holotipe yang lebih kecil memiliki berat sekitar 283 kilogram (624 pon), dengan panjang 603 meter (1.978 ft 4+1⁄4 in), dan tinggi panggul sekitar 136 meter (446 ft 2+1⁄4 in), serta panjang tengkoraknya adalah 523 milimeter (1 ft 8+1⁄2 in).[24][26] Jejak beristirahat dari teropoda yang mirip dengan Dilophosaurus dan Liliensternus telah diinterpretasikan oleh beberapa peneliti sebagai bukti cetakan bulu di sekitar perut dan kaki, yang mirip dengan bulu halus.[27][28] Namun, peneliti lain menginterpretasikan cetakan ini sebagai artefak sedimentologis yang tercipta saat dinosaurus tersebut bergerak, meskipun interpretasi ini tidak mengesampingkan kemungkinan bahwa pembuat jejak tersebut mungkin saja memiliki bulu.[29][30]
Tengkorak
sunting





Tengkorak Dilophosaurus berukuran besar jika dibandingkan secara proporsional dengan keseluruhan kerangkanya, namun rapuh. Moncongnya sempit jika dilihat dari depan, menjadi semakin sempit ke arah bagian atasnya yang membulat. Tulang pramaksila (tulang depan rahang atas) panjang dan rendah jika dilihat dari samping, menggembung di bagian depan, dan permukaan luarnya menjadi kurang cembung dari moncong hingga ke naris (lubang hidung bertulang). Lubang hidungnya terletak lebih jauh ke belakang daripada sebagian besar teropoda lainnya. Tulang-tulang pramaksila berartikulasi rapat satu sama lain, dan meskipun pramaksila hanya terhubung dengan maksila (tulang rahang atas berikutnya) di bagian tengah langit-langit mulut, tanpa koneksi di bagian samping, tulang-tulang ini membentuk sendi yang kuat melalui artikulasi kokoh yang saling mengunci antara prosesus-prosesus yang mengarah ke belakang dan ke depan dari tulang-tulang ini. Ke arah belakang dan bawah, pramaksila membentuk dinding untuk sebuah celah di antara tulang itu sendiri dan maksila yang disebut celah subnarial (juga diistilahkan sebagai "lekukan"). Celah semacam ini juga terdapat pada coelophysoid, serta dinosaurus lainnya. Celah subnarial menghasilkan sebuah diastema, yaitu sebuah celah pada deretan gigi (yang juga disebut sebagai "takik"). Di dalam celah subnarial terdapat sebuah ekskavasi dalam di belakang deretan gigi pramaksila, yang disebut lubang subnarial, yang dibatasi oleh lunas pramaksila yang mengarah ke bawah.[2][24][31][7][6]
Permukaan luar pramaksila dipenuhi oleh foramina (bukaan) dengan berbagai ukuran. Bagian atas dari dua prosesus pramaksila yang memanjang ke belakang berukuran panjang dan rendah, serta membentuk sebagian besar batas atas dari naris yang memanjang. Terdapat lekukan ke arah depan, yang membuat area di dekat dasarnya berbentuk cekung saat dilihat dari profil samping. Bagian bawah pramaksila yang berisi alveoli (soket gigi) berbentuk oval. Maksila tersebut dangkal, dan tertekan di sekitar fenestra antorbital (bukaan besar di depan mata), membentuk relung yang membulat ke arah depan, dan lebih halus daripada bagian maksila lainnya. Sebuah foramen yang disebut fenestra preanteorbital bermuara di relung ini pada lengkungan depannya. Foramina berukuran besar membentang di sisi maksila, di atas alveoli. Sebuah alur nutrien yang dalam membentang ke belakang dari lubang subnarial di sepanjang dasar lempeng interdental (atau rugosae) pada maksila.[2]

Dilophosaurus memiliki sepasang jambul yang tinggi, tipis, dan melengkung (atau berbentuk lempeng) secara longitudinal di atap tengkoraknya. Jambul ini (yang disebut jambul nasolakrimal) berawal sebagai punggungan rendah pada pramaksila dan sebagian besar dibentuk oleh tulang hidung dan tulang air mata yang meluas ke atas. Tulang-tulang ini terko-osifikasi bersama (fusi selama pembentukan jaringan tulang), sehingga letak sutura di antara keduanya tidak dapat ditentukan. Tulang lakrimal meluas menjadi tonjolan preorbital yang tebal dan kasar, membentuk lengkungan di batas depan atas orbit (rongga mata), dan menyokong bagian bawah di belakang jambul. Sebagai ciri unik bagi genus ini, tepian di atas orbit berlanjut ke belakang dan berakhir pada prosesus kecil yang hampir berbentuk segitiga di belakang orbit, yang sedikit melengkung ke arah luar. Karena hanya sebagian kecil permukaan atas dari prosesus ini yang tidak rusak, sisa jambul mungkin telah menjulang di atas tengkorak sejauh ~12 milimeter (0,47 in). Bagian jambul yang terawetkan pada UCMP 77270 adalah yang tertinggi di sekitar titik tengah panjang fenestra antorbital. UCMP 77270 mengawetkan landasan cekung di antara dasar-dasar jambul tersebut, dan apabila dilihat dari depan, jambul-jambul itu memproyeksi ke atas dan ke samping pada sudut ~80°. Welles menganggap jambul ini mengingatkan pada kasuari berjambul ganda, sementara Marsh dan Rowe menyatakan bahwa jambul tersebut kemungkinan besar ditutupi oleh keratin atau kulit yang terkeratinisasi. Mereka menunjukkan bahwa dengan membandingkannya dengan ayam-mutiara biasa, keratin pada jambul Dilophosaurus dapat memperbesar ukurannya jauh melebihi apa yang diindikasikan oleh tulangnya. Karena hanya satu spesimen yang mengawetkan sebagian besar jambul ini, tidak diketahui apakah bentuknya berbeda-beda antarindividu.[6][2][7][11][5][13] Pemindaian CT scan menunjukkan bahwa kantung udara (kantong-kantong udara yang memberikan kekuatan dan meringankan tulang) hadir di tulang-tulang yang mengelilingi otak, dan bersambungan dengan rongga sinus di bagian depan tengkorak. Fenestra antorbital bersambungan dengan sisi jambul, yang mengindikasikan bahwa jambul tersebut juga memiliki kantung udara (sebuah punggungan tulang membentuk atap di atas fenestra antorbital pada sebagian besar teropoda lainnya).[15]

Orbitnya berbentuk oval, dan menyempit ke arah bawah. Tulang jugal memiliki dua prosesus yang mengarah ke atas, yang pertama membentuk bagian tepi bawah dari fenestra antorbital, dan bagian margin bawah dari orbit. Sebuah proyeksi dari tulang kuadrat ke dalam fenestra temporal lateral (bukaan di belakang mata) memberikannya garis luar yang berbentuk seperti ginjal (reniform). Foramen magnum (bukaan besar di bagian belakang tempurung otak) lebarnya sekitar setengah dari kondilus oksipital, yang mana kondilus itu sendiri berbentuk seperti hati (kordiform), serta memiliki leher yang pendek dan alur di bagian sampingnya.[2] Tulang rahang bawah berbentuk ramping dan tipis di bagian depan, tetapi regio artikularnya (tempat ia terhubung dengan tengkorak) berukuran masif, dan rahang bawahnya menebal di sekitar fenestra mandibula (sebuah bukaan di bagian sampingnya). Fenestra mandibula berukuran kecil pada Dilophosaurus, dibandingkan dengan milik kelompok coelophysoid, dan menyusut dari depan ke belakang, sebuah keunikan tersendiri bagi genus ini. Tulang dentari (bagian depan rahang bawah tempat sebagian besar gigi menempel) memiliki dagu yang melengkung ke atas alih-alih meruncing. Dagu ini memiliki foramen yang besar di ujungnya, dan sederet foramina kecil membentang secara kasar sejajar dengan tepi atas dentari. Di sisi dalam, simfisis mandibula (tempat dua bagian rahang bawah terhubung) berbentuk rata dan halus, serta tidak menunjukkan tanda-tanda menyatu dengan bagian pasangannya. Sebuah foramen Meckel membentang di sepanjang sisi luar dentari. Permukaan samping dari tulang surangular memiliki prosesus piramidal yang unik di depan artikulasi dengan kuadrat, dan punggung bukit horizontal ini membentuk sebuah landasan. Prosesus retroartikular pada rahang bawah (sebuah proyeksi yang mengarah ke belakang) berukuran panjang.[2][31][6]
Dilophosaurus memiliki empat gigi di setiap pramaksila, 12 di setiap maksila, dan 17 di setiap dentari. Gigi-giginya umumnya berukuran panjang, tipis, dan melengkung, dengan dasar yang relatif kecil. Gigi-gigi ini termampatkan di bagian samping, penampang lintang di dasarnya berbentuk oval, lentikular (berbentuk lensa) di bagian atas, dan sedikit cekung di sisi luar serta dalamnya. Gigi maksila terbesar berada di dalam atau di dekat alveolus keempat, dan tinggi mahkota giginya semakin mengecil ke arah belakang. Gigi pertama pada maksila sedikit mengarah ke depan dari alveolusnya karena batas bawah prosesus pramaksila (yang memproyeksi ke belakang ke arah maksila) melengkung ke atas. Gigi-gigi dentari jauh lebih kecil daripada gigi-gigi maksila. Gigi ketiga atau keempat pada dentari Dilophosaurus dan beberapa coelophysoid adalah yang terbesar di bagian tersebut, dan sepertinya pas untuk masuk ke dalam celah subnarial di rahang atas. Sebagian besar gigi memiliki gerigi di tepi depan dan belakangnya, yang diimbangi oleh alur-alur vertikal, dan berukuran lebih kecil di bagian depan. Terdapat sekitar 31 hingga 41 gerigi di tepi depan, dan 29 hingga 33 di tepi belakang. Setidaknya gigi kedua dan ketiga pada pramaksila memiliki gerigi, tetapi gigi keempat tidak. Gigi-gigi ini ditutupi oleh lapisan tipis email, dengan ketebalan 01 hingga 015 mm (0,039 hingga 0,591 in), yang meluas jauh hingga ke dasarnya. Alveolinya berbentuk elips hingga hampir melingkar, dan semuanya lebih besar daripada dasar gigi yang dikandungnya, yang karenanya gigi-gigi ini mungkin tertanam secara longgar di rahang. Meskipun jumlah alveoli pada dentari tampaknya mengindikasikan bahwa giginya sangat berdesakan, jarak antar giginya agak berjauhan, karena ukuran alveolinya yang lebih besar. Rahangnya berisi gigi pengganti pada berbagai tahap erupsi. Lempeng interdental di antara gigi-giginya sangat rendah.[2][31][12]
Kerangka pascakranial
sunting





Dilophosaurus memiliki 10 tulang belakang servikal (leher), 14 dorsal (punggung), dan 45 kaudal (ekor), dan kantung udara tumbuh ke dalam tulang-tulang belakang tersebut. Lehernya panjang, yang kemungkinan dapat ditekuk hampir 90° oleh tengkorak dan bahunya, menahan tengkoraknya dalam postur horizontal. Tulang belakang servikalnya sangat ringan; bagian sentranya ("badan" dari tulang belakang) dilubangi oleh pleurosoel (cekungan di bagian samping) dan sentrosoel (rongga di bagian dalam). Lengkungan tulang belakang servikal juga memiliki fosa pneumatik (atau konosis), yaitu relung berbentuk kerucut yang sangat besar sehingga tulang yang memisahkannya terkadang setipis kertas. Sentranya bersifat plano-konkaf, yakni datar hingga sedikit cembung di bagian depan dan mencekung dalam di bagian belakang, mirip dengan Ceratosaurus. Hal ini mengindikasikan bahwa lehernya fleksibel, meskipun ia memiliki tulang rusuk leher yang panjang, tumpang tindih, dan menyatu dengan sentra. Tulang rusuk lehernya ramping dan mungkin mudah ditekuk.[2][31][6][15]
Tulang atlas (tulang belakang servikal pertama yang menempel pada tengkorak) memiliki sentrum kecil berbentuk kubus, dan memiliki cekungan di bagian depan tempat tulang ini membentuk mangkuk untuk kondilus oksipital (tonjolan yang terhubung dengan tulang belakang atlas) di bagian belakang tengkorak. Tulang aksis (tulang belakang servikal kedua) memiliki duri saraf yang berat, dan postzigapofisisnya (prosesus tulang belakang yang berartikulasi dengan prezigapofisis dari tulang belakang berikutnya) bertemu dengan prezigapofisis panjang yang melengkung ke atas dari tulang belakang servikal ketiga. Sentra dan duri saraf tulang belakang servikal berukuran panjang dan rendah, serta duri-durinya berundak jika dilihat dari samping, membentuk "bahu" di bagian depan dan belakang, serta "topi" tengah yang lebih tinggi yang memberikan tampilan seperti Salib Malta (krusiform) jika dilihat dari atas, yang mana ini merupakan ciri khas dinosaurus tersebut. Lamina sentrodiapofiseal posterior pada tulang servikal menunjukkan variasi serial, bercabang dua dan menyatu kembali di sepanjang leher, sebuah ciri yang unik. Duri saraf dari tulang belakang dorsal juga rendah serta meluas di depan dan belakang, yang membentuk perlekatan kuat untuk ligamen. Sebagai keunikan genus ini, terdapat tambahan lamina yang memancar dari lamina sentrodiapofiseal anterior dan lamina sentrodiapofiseal posterior pada tulang belakang punggung tengah. Tulang belakang sakral yang menempati sepanjang bilah ilium tampaknya tidak menyatu. Tulang rusuk dari tulang belakang sakral pertama berartikulasi dengan prosesus preasetabular pada ilium, sebuah ciri yang khas. Sentra pada tulang belakang kaudal sangat konsisten dalam hal panjang, tetapi diameternya menjadi semakin kecil ke arah belakang, dan penampang lintangnya berubah dari elips menjadi melingkar.[2][31][6]






Skapula (tulang belikat) berukuran sedang panjangnya dan cekung di sisi dalamnya untuk mengikuti kelengkungan tubuh. Skapula ini lebar, terutama bagian atasnya, yang berbentuk persegi panjang (atau bersudut siku), sebuah ciri yang unik. Korakoid berbentuk elips, dan tidak menyatu dengan skapula. Bagian belakang bawah dari korakoid memiliki "penyangga horizontal" di sebelah tuber bisep, unik untuk genus ini. Lengannya kuat, dan memiliki lubang-lubang yang dalam serta prosesus yang kokoh untuk perlekatan otot dan ligamen. Humerus (tulang lengan atas) berukuran besar dan ramping, dan ulna (tulang lengan bawah) berbentuk kokoh dan lurus, dengan olekranon yang kokoh. Tangannya memiliki empat jari: jari pertama lebih pendek tetapi lebih kuat daripada dua jari berikutnya, dengan sebuah cakar besar, dan dua jari berikutnya lebih panjang dan ramping, dengan cakar yang lebih kecil. Cakar-cakarnya melengkung dan tajam. Jari ketiganya mereduksi, dan jari keempat bersifat vestigial (dipertahankan, tetapi tidak memiliki fungsi).[2][31][6]
Krista ilium berada di posisi tertinggi di atas pedunkel ilium (prosesus ilium yang mengarah ke bawah), dan sisi luarnya berbentuk cekung. Bagian kaki dari tulang pubis hanya sedikit meluas, sedangkan ujung bawahnya meluas jauh lebih besar pada iskium, yang juga memiliki batang tulang yang sangat tipis. Tungkai belakangnya besar, dengan femur (tulang paha) yang sedikit lebih panjang daripada tibia (tulang betis bagian dalam), yang mana ini merupakan kebalikan dari, misalnya, Coelophysis. Femurnya masif; batang tulangnya berbentuk sigmoid (melengkung seperti huruf 'S'), dan trokanter mayornya terpusat pada batang tersebut. Tibianya memiliki tuberositas yang berkembang baik dan membesar di ujung bawah. Astragalus (tulang pergelangan kaki) terpisah dari tibia dan kalkaneus, serta membentuk separuh dari soket untuk tulang fibula. Hewan ini memiliki kaki yang panjang dan kokoh dengan tiga jari kaki yang berkembang baik yang menanggung cakar-cakar besar, yang kelengkungannya jauh lebih kecil daripada cakar pada tangan. Jari kaki ketiga adalah yang paling kokoh, dan jari kaki pertama yang lebih kecil (haluks) dipertahankan agar tidak menyentuh tanah.[2][31][32][6]
Klasifikasi
sunting
Welles menganggap Dilophosaurus sebagai seekor megalosaurus pada tahun 1954, namun merevisi pendapatnya pada tahun 1970 setelah menemukan bahwa hewan ini memiliki jambul.[7][3] Pada tahun 1974, Welles dan ahli paleontologi Robert A. Long mengklasifikasikan Dilophosaurus sebagai seekor ceratosauroid.[33] Pada tahun 1984 Welles menemukan bahwa Dilophosaurus menunjukkan ciri-ciri dari Coelurosauria dan Carnosauria, dua kelompok utama yang menjadi pembagian teropoda hingga saat itu, berdasarkan ukuran tubuh, dan ia mengusulkan bahwa pembagian ini tidak akurat. Ia menemukan bahwa Dilophosaurus paling dekat kerabatnya dengan teropoda-teropoda yang biasanya ditempatkan dalam famili Halticosauridae, khususnya Liliensternus.[2]
Pada tahun 1988, ahli paleontologi Gregory S. Paul mengklasifikasikan halticosauria sebagai subfamili dari famili Coelophysidae, dan mengusulkan bahwa Dilophosaurus kemungkinan merupakan keturunan langsung dari Coelophysis. Paul juga mempertimbangkan kemungkinan bahwa spinosaurus adalah dilophosaurus yang sintas hingga masa berikutnya, berdasarkan kemiripan pada moncong yang tertekuk, posisi lubang hidung, dan gigi-gigi yang ramping pada Baryonyx.[24] Pada tahun 1994, ahli paleontologi Thomas R. Holtz menempatkan Dilophosaurus ke dalam kelompok Coelophysoidea, bersama dengan namun terpisah dari Coelophysidae. Ia menempatkan Coelophysoidea ke dalam kelompok Ceratosauria.[34] Pada tahun 2000, ahli paleontologi James H. Madsen dan Welles membagi Ceratosauria ke dalam famili Ceratosauridae dan Dilophosauridae, dengan Dilophosaurus sebagai satu-satunya anggota dari famili yang disebut terakhir.[35]

Lamanna dan rekan-rekannya pada tahun 1998 menyoroti bahwa semenjak Dilophosaurus ditemukan memiliki jambul pada tengkoraknya, teropoda berjambul serupa lainnya juga telah ditemukan (termasuk Sinosaurus), dan bahwa ciri ini, oleh karenanya, tidak unik bagi genus tersebut, serta penggunaannya terbatas untuk menentukan hubungan kekerabatan di dalam kelompok mereka.[21] Ahli paleontologi Adam M. Yates mendeskripsikan genus Dracovenator dari Afrika Selatan pada tahun 2005, dan menemukannya berkerabat dekat dengan Dilophosaurus dan Zupaysaurus. Analisis kladistiknya menunjukkan bahwa mereka tidak termasuk dalam Coelophysoidea, melainkan dalam Neotheropoda, sebuah kelompok yang lebih terturunkan (atau "maju"). Ia mengusulkan bahwa jika Dilophosaurus lebih terturunkan dari Coelophysoidea, ciri-ciri yang dimilikinya bersama dengan kelompok ini mungkin diwarisi dari teropoda yang basal (atau "primitif"), yang mengindikasikan bahwa teropoda mungkin telah melewati "tahap coelophysoid" dalam evolusi awal mereka.[36]
Pada tahun 2007, ahli paleontologi Nathan D. Smith dan rekan-rekannya menemukan teropoda berjambul Cryolophosaurus sebagai spesies saudari dari Dilophosaurus, dan mengelompokkan mereka dengan Dracovenator dan Sinosaurus. Klad ini lebih terturunkan dari Coelophysoidea, namun lebih basal dari Ceratosauria, dengan demikian menempatkan teropoda basal dalam susunan yang menyerupai tangga.[37] Pada tahun 2012, Carrano dan rekan-rekannya menemukan bahwa kelompok teropoda berjambul yang diusulkan oleh Smith dan rekan-rekannya didasarkan pada ciri-ciri yang berkaitan dengan keberadaan jambul semacam itu, namun ciri-ciri pada sisa kerangka lainnya kurang konsisten. Mereka justru menemukan bahwa Dilophosaurus merupakan bagian dari coelophysoid, dengan Cryolophosaurus dan Sinosaurus menjadi lebih terturunkan, sebagai anggota basal dari kelompok Tetanurae.[13]
Ahli paleontologi Christophe Hendrickx dan rekan-rekannya mendefinisikan Dilophosauridae untuk menyertakan Dilophosaurus dan Dracovenator pada tahun 2015, dan mencatat bahwa meskipun terdapat ketidakpastian umum mengenai penempatan kelompok ini, tampaknya kelompok ini sedikit lebih terturunkan dari Coelophysoidea, dan merupakan kelompok saudari bagi Averostra. Kelompok Dilophosauridae berbagi ciri-ciri dengan Coelophysoidea seperti celah subnarial dan gigi-gigi depan pada maksila yang mengarah ke depan, sedangkan ciri-ciri yang dimilikinya bersama Averostra mencakup sebuah fenestra di bagian depan maksila dan berkurangnya jumlah gigi pada maksila. Mereka menyarankan bahwa jambul kranial pada Cryolophosaurus dan Sinosaurus dapat saja telah berevolusi secara konvergen, atau merupakan ciri yang diwarisi dari nenek moyang yang sama. Kladogram berikut ini didasarkan pada apa yang dipublikasikan oleh Hendrickx dan rekan-rekannya, yang mana juga didasarkan pada penelitian-penelitian sebelumnya:[38]

Pada tahun 2019, ahli paleontologi Marion Zahner dan Winand Brinkmann menemukan bahwa anggota Dilophosauridae merupakan taksa saudari basal yang berturut-turut dari Averostra, alih-alih sebuah klad monofiletik (kelompok alami), tetapi mencatat bahwa beberapa dari analisis mereka mendapati kelompok tersebut valid, yang mengandung Dilophosaurus, Dracovenator, Cryolophosaurus, dan kemungkinan Notatesseraeraptor sebagai anggota paling basal. Oleh karena itu, mereka memberikan diagnosis untuk Dilophosauridae, berdasarkan ciri-ciri pada rahang bawah.[39] Dalam analisis filogenetik yang menyertai pendeskripsian ulang mereka pada tahun 2020, Marsh dan Rowe menemukan bahwa seluruh spesimen Dilophosaurus membentuk kelompok monofiletik, yang bersaudari dengan Averostra, dan lebih terturunkan dari Cryolophosaurus. Analisis mereka tidak menemukan dukungan untuk Dilophosauridae, dan mereka menyarankan bahwa jambul kranial merupakan sifat plesiomorfik (leluhur) dari Ceratosauria dan Tetanurae.[6]
Iknologi
sunting

Berbagai iknotaksa (taksa yang didasarkan pada fosil jejak) telah dikaitkan dengan Dilophosaurus atau teropoda serupa. Pada tahun 1971, Welles melaporkan jejak kaki dinosaurus dari Formasi Kayenta di Arizona utara, pada dua lapisan yakni 14 m (45 ft) dan 112 m (367 ft) di bawah lokasi ditemukannya spesimen asli Dilophosaurus. Jejak kaki di lapisan bawah bersifat tridaktil (berjari tiga), dan bisa saja dibuat oleh Dilophosaurus; Welles mendirikan iknogenus dan spesies baru Dilophosauripus williamsi berdasarkan temuan tersebut, untuk menghormati Williams, penemu kerangka Dilophosaurus pertama. Spesimen tipenya adalah cetakan jejak kaki besar yang dikatalogkan sebagai UCMP 79690-4, bersama cetakan dari tiga jejak lain yang disertakan dalam hipodigma.[40] Pada tahun 1984, Welles mengakui bahwa belum ada cara yang ditemukan untuk membuktikan atau menyangkal apakah jejak kaki tersebut memang milik Dilophosaurus.[2] Pada tahun 1996, ahli paleontologi Michael Morales dan Scott Bulkey melaporkan sebuah lintasan jejak dari iknogenus Eubrontes dari Formasi Kayenta yang dibuat oleh seekor teropoda yang sangat besar. Mereka mencatat bahwa jejak itu mungkin dibuat oleh individu Dilophosaurus yang sangat besar, namun menganggap hal itu tidak mungkin, karena mereka memperkirakan pembuat jejak tersebut memiliki tinggi 283–299 m (928 ft 5+3⁄4 in – 980 ft 11+3⁄4 in) di panggul, dibandingkan dengan Dilophosaurus yang tingginya 150–175 m (492 ft 1+1⁄2 in – 574 ft 1+3⁄4 in).[41]
Ahli paleontologi Gerard Gierliński memeriksa jejak kaki tridaktil dari Pegunungan Salib Suci di Polandia dan menyimpulkan pada tahun 1991 bahwa jejak-jejak itu milik teropoda seperti Dilophosaurus. Ia menamai iknospesies baru Grallator (Eubrontes) soltykovensis berdasarkan jejak-jejak tersebut, dengan cetakan jejak kaki MGIW 1560.11.12 sebagai holotipe.[42] Pada tahun 1994, Gierliński juga mengaitkan jejak kaki dari Formasi Höganäs di Swedia yang ditemukan pada tahun 1974 ke G. (E.) soltykovensis.[43] Pada tahun 1996, Gierliński mengaitkan jejak AC 1/7 dari Formasi Turners Falls di Massachusetts, sebuah jejak istirahat yang ia yakini menunjukkan cetakan bulu, dengan teropoda yang mirip dengan Dilophosaurus dan Liliensternus, dan memasukkannya ke dalam iknotakson Grallator minisculus.[27] Ahli paleontologi Martin Kundrát pada tahun 2004 sepakat bahwa jejak tersebut menunjukkan cetakan bulu, namun interpretasi ini dibantah oleh ahli paleontologi Martin Lockley dan rekan-rekannya pada tahun 2003 serta ahli paleontologi Anthony J. Martin dan rekan-rekannya pada tahun 2004, yang menganggapnya sebagai artefak sedimentologis. Martin dan rekan-rekannya juga memindahkan jejak tersebut ke dalam iknotakson Fulicopus lyellii.[28][29][30] Gierliński dan Karol Sabath merespons dalam sebuah presentasi konferensi pada tahun 2005, dengan menyoroti bahwa cetakan hamparan alga tidak hanya akan terbentuk pada bagian perut, tetapi juga pada jejak kakinya. Berdasarkan eksperimen dan foto-foto yang mendetail, mereka menemukan bahwa jejak tersebut mirip dengan jejak yang ditinggalkan oleh bulu berserat (semiplum) dari burung modern, dan berbeda dari jejak yang ditinggalkan oleh tubuh bersisik.[44]

Ahli paleontologi Robert E. Weems mengusulkan pada tahun 2003 bahwa jejak Eubrontes tidak dibuat oleh seekor teropoda, melainkan oleh sauropodomorfa yang mirip dengan Plateosaurus, sehingga menyingkirkan Dilophosaurus sebagai kandidat pembuat jejaknya. Sebaliknya, Weems mengusulkan Kayentapus hopii, iknotakson lain yang dinamai oleh Welles pada tahun 1971, sebagai pasangan yang paling cocok untuk Dilophosaurus.[45] Kaitannya dengan Dilophosaurus terutama didasarkan pada sudut lebar antara cetakan jari ketiga dan keempat yang ditunjukkan oleh jejak-jejak ini, dan pengamatan bahwa kaki spesimen holotipe menunjukkan jari keempat yang merentang ke luar dengan cara yang serupa. Juga pada tahun 2003, ahli paleontologi Emma Rainforth berargumen bahwa rentangan pada kaki holotipe tersebut hanyalah akibat distorsi, dan bahwa Eubrontes memang merupakan pasangan yang cocok untuk Dilophosaurus.[46][47] Ahli paleontologi Spencer G. Lucas dan rekan-rekannya menyatakan pada tahun 2006 bahwa nyaris terdapat kesepakatan universal bahwa jejak Eubrontes dibuat oleh teropoda seperti Dilophosaurus, dan bahwa mereka beserta para peneliti lainnya menolak klaim Weems.[48]
Pada tahun 2006, Weems mempertahankan penilaian tahun 2003-nya mengenai Eubrontes, dan mengusulkan hewan seperti Dilophosaurus sebagai pembuat jejak yang mungkin untuk berbagai lintasan jejak Kayentapus di Tambang Culpeper di Virginia. Weems menyarankan bahwa cetakan membulat yang berasosiasi dengan beberapa lintasan jejak ini merepresentasikan cetakan tangan yang tidak memiliki jejak jari, yang ia interpretasikan sebagai jejak pergerakan kuadrupedal (berkaki empat).[47] Milner dan rekan-rekannya menggunakan kombinasi baru Kayentapus soltykovensis pada tahun 2009, dan menyarankan bahwa Dilophosauripus mungkin tidak berbeda secara definitif dari Eubrontes dan Kayentapus. Mereka mengusulkan bahwa bekas cakar panjang yang digunakan untuk membedakan Dilophosauripus mungkin merupakan sebuah artefak akibat seretan. Mereka menemukan bahwa jejak Gigandipus dan Anchisauripus mungkin juga hanya merepresentasikan variasi dari Eubrontes. Mereka menyoroti bahwa perbedaan di antara iknotaksa mungkin lebih mencerminkan cara pembuat jejak berinteraksi dengan substrat ketimbang perbedaan taksonomi. Mereka juga menemukan bahwa Dilophosaurus merupakan pasangan yang cocok untuk sebuah lintasan jejak dan jejak istirahat Eubrontes (SGDS 18.T1) dari Situs Penemuan Dinosaurus St. George di Formasi Moenave di Utah, meskipun dinosaurus itu sendiri tidak diketahui dari formasi tersebut, yang mana usianya sedikit lebih tua dari Formasi Kayenta.[49]
Weems menyatakan pada tahun 2019 bahwa jejak Eubrontes tidak mencerminkan kaki ramping Dilophosaurus, dan berargumen bahwa jejak tersebut malah dibuat oleh sauropodomorfa bipedal Anchisaurus.[50] Dalam tinjauan jejak masa Jura pada tahun 2024, ahli paleontologi John R. Foster dan rekan-rekannya menyatakan bahwa sangat sedikit iknolog lain yang menerima interpretasi sauropodomorfa dari Weems mengenai Eubrontes, sebagian karena jejak-jejak semacam itu melimpah di tempat-tempat di mana tidak ada fosil sauropodomorfa yang ditemukan.[51]
Paleobiologi
suntingPola makan dan diet
sunting
Welles menemukan bahwa Dilophosaurus tidak memiliki gigitan yang kuat, karena kelemahan yang disebabkan oleh celah subnarial. Ia berpendapat bahwa hewan ini menggunakan gigi pramaksila depannya untuk memetik dan mencabik alih-alih menggigit, dan gigi maksila di bagian belakang untuk menusuk dan mengiris. Ia menduga bahwa hewan ini kemungkinan adalah pemakan bangkai alih-alih pemangsa, dan jika ia memang membunuh hewan besar, hal itu dilakukannya menggunakan tangan dan kakinya ketimbang rahangnya. Welles tidak menemukan bukti adanya kinesis kranial pada tengkorak Dilophosaurus, sebuah ciri yang memungkinkan tulang-tulang penyusun tengkorak untuk bergerak satu sama lain.[2] Pada tahun 1986, ahli paleontologi Robert T. Bakker sebaliknya menemukan bahwa Dilophosaurus, dengan leher dan tengkoraknya yang masif serta gigi atasnya yang besar, telah beradaptasi untuk membunuh mangsa berukuran besar, dan cukup kuat untuk menyerang herbivora Jura Awal mana pun.[52] Pada tahun 1988, Paul menepis gagasan bahwa Dilophosaurus adalah pemakan bangkai, dan mengklaim bahwa hewan darat yang sepenuhnya pemakan bangkai adalah mitos. Ia menyatakan bahwa moncong Dilophosaurus tertopang lebih baik dari yang dikira sebelumnya, dan bahwa gigi maksilanya yang sangat besar dan ramping lebih mematikan daripada cakarnya. Paul mengusulkan bahwa ia memburu hewan besar seperti prosauropoda, dan bahwa ia lebih mampu mencaplok hewan-hewan kecil dibandingkan teropoda lain yang berukuran serupa.[24]
Sebuah studi teori balok pada tahun 2005 oleh ahli paleontologi François Therrien dan rekan-rekannya menemukan bahwa kekuatan gigitan pada rahang bawah Dilophosaurus menurun drastis ke arah belakang di deretan giginya. Hal ini mengindikasikan bahwa bagian depan rahang bawah, dengan dagunya yang melengkung ke atas, "roset" giginya, dan wilayah simfisis yang diperkuat (mirip dengan spinosaurid), digunakan untuk menangkap dan memanipulasi mangsa, yang kemungkinan berukuran relatif lebih kecil. Sifat-sifat simfisis mandibulanya mirip dengan felid (kucing-kucingan) dan buaya yang menggunakan bagian depan rahangnya untuk memberikan gigitan yang kuat saat menaklukkan mangsa. Beban yang diberikan pada rahang bawah konsisten dengan perlawanan mangsa berukuran kecil, yang mungkin diburu dengan memberikan gigitan menyayat untuk melukainya, dan kemudian ditangkap menggunakan bagian depan rahang setelah mangsa tersebut terlalu lemah untuk melawan. Mangsa tersebut lalu mungkin dipindahkan lebih jauh ke belakang di dalam rahang, tempat letak gigi-gigi terbesar, dan dibunuh melalui gigitan mengiris (mirip dengan beberapa jenis buaya) menggunakan gigi-giginya yang memipih di bagian samping. Para penulis studi tersebut menyarankan bahwa jika Dilophosaurus memang memangsa hewan-hewan kecil, kemungkinan kawanannya berburu dalam jumlah yang terbatas.[53]
Milner dan ahli paleontologi James I. Kirkland pada tahun 2007 menyarankan bahwa Dilophosaurus memiliki ciri-ciri yang mengindikasikan bahwa ia mungkin memakan ikan. Mereka menyoroti bahwa ujung rahangnya melebar ke samping, membentuk "roset" berupa gigi yang saling mengunci, mirip dengan spinosaurid yang diketahui memakan ikan, dan gharial, yakni buaya modern yang paling banyak memakan ikan. Bukaan hidungnya juga tertarik ke belakang pada rahang, mirip dengan spinosaurid, yang memiliki bukaan hidung yang lebih tertarik lagi, dan hal ini mungkin telah membatasi masuknya percikan air ke dalam lubang hidung selama menangkap ikan. Kedua kelompok tersebut juga memiliki lengan yang panjang dengan cakar yang berkembang baik, yang dapat membantu saat menangkap ikan. Danau Dixie, sebuah danau besar yang membentang dari Utah ke Arizona dan Nevada, kemungkinan menyediakan banyak ikan di dunia "pasca-bencana" yang secara biologis lebih miskin menyusul terjadinya peristiwa kepunahan Trias–Jura (di mana sekitar tiga perempat kehidupan di Bumi musnah), 5 hingga 15 juta tahun sebelum Dilophosaurus muncul.[54][15]

Pada tahun 2018, Marsh dan Rowe melaporkan bahwa spesimen holotipe sauropodomorfa Sarahsaurus memiliki kemungkinan bekas gigi yang tersebar di seluruh kerangkanya yang mungkin ditinggalkan oleh Dilophosaurus (Syntarsus terlalu kecil untuk membuatnya) yang memakan bangkai spesimen tersebut setelah mati (posisi tulang-tulangnya mungkin juga telah terganggu akibat aktivitas memakan bangkai tersebut). Salah satu contoh bekas tersebut dapat dilihat pada skapula kiri, yang memiliki cekungan berbentuk oval di permukaan sisi atasnya, dan sebuah lubang besar di ujung depan bawah dari tibia kanannya. Tambang tempat digalinya spesimen holotipe dan paratipe Sarahsaurus juga menyimpan spesimen Dilophosaurus remaja yang tidak lengkap.[55] Marsh dan Rowe menyarankan pada tahun 2020 bahwa banyak dari ciri yang membedakan Dilophosaurus dengan teropoda yang lebih awal berkaitan dengan pertambahan ukuran tubuh dan makropredasi (memangsa hewan besar). Meskipun Marsh dan Rowe sepakat bahwa Dilophosaurus bisa saja memakan ikan dan mangsa kecil di sistem sungai pada lingkungannya, mereka menyoroti bahwa artikulasi antara pramaksila dan maksila pada rahang atas bersifat tidak dapat digerakkan dan jauh lebih kokoh dari yang dikira sebelumnya, dan mangsa bertubuh besar bisa saja dicengkeram serta dimanipulasi dengan tungkai depan selama proses memangsa dan memakan bangkai. Mereka menganggap bekas gigitan besar pada spesimen Sarahsaurus bersama dengan gigi yang tanggal serta keberadaan spesimen Dilophosaurus di dalam tambang yang sama sebagai dukungan untuk gagasan ini.[6]
Dalam sebuah artikel tahun 2021, ahli paleontologi Matthew A. Brown dan Rowe menyatakan bahwa sisa-sisa fosil ini menunjukkan bahwa Dilophosaurus memiliki rahang yang cukup kuat untuk menembus tulang. Kantung udara berdaging dari sistem pernapasannya yang tumbuh ke dalam tulang belakang tidak hanya memperkuat melainkan juga meringankan kerangkanya, serta memungkinkan aliran udara satu arah melalui paru-parunya, mirip dengan burung dan buaya, dan dengan demikian menghasilkan lebih banyak oksigen daripada sistem pernapasan dua arah pada mamalia (di mana udara mengalir masuk dan keluar dari paru-paru). Pernapasan satu arah mengindikasikan tingkat metabolisme yang relatif tinggi dan karenanya memengaruhi tingginya tingkat aktivitas, yang menandakan bahwa Dilophosaurus kemungkinan adalah pemburu yang cepat dan tangkas. Brown dan Rowe menganggap Dilophosaurus sebagai predator puncak di ekosistemnya, dan bukan pemakan bangkai.[15]
Pergerakan
sunting
Welles membayangkan Dilophosaurus sebagai hewan bipedal yang jelas-jelas aktif, mirip dengan burung unta yang diperbesar. Ia menemukan bahwa tungkai depannya merupakan senjata yang ampuh, kuat, serta fleksibel, dan tidak digunakan untuk bergerak. Ia mencatat bahwa tangannya mampu mencengkeram dan menyayat, saling bersentuhan satu sama lain, dan menjangkau hingga dua pertiga bagian lehernya. Ia mengusulkan bahwa dalam postur duduk, hewan ini akan bersandar pada "kaki" besar dari iskiumnya, di samping mengandalkan ekor dan kaki bawahnya.[2] Pada tahun 1990, ahli paleontologi Stephen dan Sylvia Czerkas menyarankan bahwa panggul Dilophosaurus yang lemah bisa saja merupakan bentuk adaptasi untuk gaya hidup akuatik, di mana air akan membantu menopang berat badannya, dan hewan ini bisa jadi merupakan perenang yang efisien. Walaupun begitu, mereka meragukan bahwa hewan ini hanya terbatas pada lingkungan perairan saja, karena melihat kekuatan dan proporsi tungkai belakangnya, yang memungkinkannya bergerak cepat dan tangkas selama pergerakan bipedalnya.[56] Pada tahun 1998, ahli paleontologi Per Christiansen memperkirakan kecepatan tertinggi Dilophosaurus mencapai 43–69 km (27–43 mi) per jam.[57] Paul menggambarkan Dilophosaurus pada tahun 2000 memantul menggunakan ekornya sembari menyerang musuh, mirip dengan seekor kanguru.[58]
Pada tahun 2005, ahli paleontologi Phil Senter dan James H. Robins memeriksa rentang gerak pada tungkai depan Dilophosaurus dan teropoda lainnya. Mereka menemukan bahwa Dilophosaurus mampu menarik humerus-nya ke belakang hingga nyaris sejajar dengan skapulanya, namun tidak dapat menggerakkannya ke depan lebih dari orientasi vertikal. Sikunya dapat mendekati ekstensi (rentangan penuh) dan fleksi (tekukan) pada sudut siku-siku, tetapi tidak dapat mencapainya secara seutuhnya. Jari-jarinya tampaknya tidak dapat dihiperekstensikan (mampu merentang ke belakang melampaui rentang normalnya) secara sukarela, tetapi jari-jari tersebut mungkin saja dapat dihiperekstensikan secara pasif untuk menahan terjadinya dislokasi selama pergerakan kasar oleh mangsa yang tertangkap.[59] Sebuah artikel tahun 2015 oleh Senter dan Robins memberikan rekomendasi tentang cara merekonstruksi postur tungkai depan pada dinosaurus bipedal, berdasarkan pemeriksaan pada berbagai taksa, termasuk Dilophosaurus. Skapulanya dipertahankan secara sangat horizontal, orientasi siku saat beristirahat akan mendekati sudut siku-siku, dan orientasi tangannya tidak akan banyak menyimpang dari orientasi lengan bawah.[60]

Pada tahun 2018, Senter dan Corwin Sullivan memeriksa rentang gerak persendian tungkai depan Dilophosaurus dengan memanipulasi tulang-tulangnya, untuk menguji hipotesis fungsi tungkai depannya. Mereka juga mempertimbangkan bahwa eksperimen yang dilakukan pada bangkai aligator menunjukkan bahwa rentang pergerakan pada siku yang tertutupi jaringan lunak (seperti tulang rawan, ligamen, dan otot) jauh lebih besar daripada apa yang diindikasikan oleh manipulasi tulang-tulang telanjangnya. Mereka menemukan bahwa humerus Dilophosaurus dapat ditarik ke posisi yang hampir sejajar dengan skapula, ditarik ke depan hingga tingkat yang nyaris vertikal, dan diangkat setinggi 65°. Siku tidak dapat ditekuk melewati sudut siku-siku terhadap humerus. Pronasi dan supinasi pergelangan tangan (menyilangkan tulang radius dan ulna pada lengan bawah untuk memutar tangan) terhalang oleh persendian radius dan ulna yang tidak dapat memutar, dan karenanya telapak tangannya menghadap ke tengah (medial), saling berhadapan satu sama lain. Ketidakmampuan mempronasi pergelangan tangan merupakan ciri leluhur yang dimiliki oleh teropoda dan kelompok dinosaurus lainnya. Pergelangan tangannya memiliki mobilitas terbatas, dan jari-jarinya memisah selama fleksi, serta sangat dapat dihiperekstensikan.[61]
Senter dan Sullivan menyimpulkan bahwa Dilophosaurus mampu menggenggam dan menahan objek di antara kedua tangan, menggenggam dan menahan objek kecil dengan satu tangan, meringkus objek di dekat bawah dadanya, membawa objek ke mulutnya, melakukan pertunjukan pamer dengan mengayunkan lengan membentuk lengkungan di sepanjang sisi tulang rusuknya, menggaruk dada, perut, atau separuh dari tungkai depan lainnya yang terjauh dari tubuh, menangkap mangsa di bawah dada atau pangkal leher, dan mendekap objek ke dadanya. Dilophosaurus tidak mampu melakukan penggalian garukan, tarikan kait, menjepit benda di antara dua ujung jari pada satu tangan, menjaga keseimbangan dengan merentangkan lengannya ke arah samping, atau menyelidiki celah-celah kecil seperti yang dilakukan oleh aye-aye modern. Kemampuan hiperekstensi pada jari-jari ini mungkin telah mencegah terjadinya dislokasi akibat perlawanan sengit dari mangsanya, lantaran hal ini memungkinkan gerak jari yang lebih besar (tanpa memengaruhi pergerakan lokomosi). Mobilitas bahu yang terbatas dan pendeknya tungkai depan mengindikasikan bahwa mulutnyalah yang pertama kali melakukan sentuhan dengan mangsa, bukan tangannya. Penangkapan mangsa dengan tungkai depannya hanya dimungkinkan untuk menangkap hewan-hewan yang cukup kecil agar muat di bawah dada Dilophosaurus, atau mangsa berukuran lebih besar yang telah dipaksa jatuh ke tanah menggunakan mulutnya. Panjang kepala dan lehernya yang luar biasa akan memungkinkan moncongnya menjangkau jauh lebih panjang dari tangannya.[61]


Jejak kaki Dilophosauripus yang dilaporkan oleh Welles pada tahun 1971 semuanya berada di lapisan yang sama, dan dideskripsikan sebagai jejak kaki yang "tumpang-tindih berantakan layaknya di halaman ayam", dengan sedikit di antaranya yang membentuk lintasan jejak. Jejak-jejak tersebut tercetak di atas lumpur, yang memungkinkan kaki-kaki tersebut tenggelam sedalam 5–10 cm (2–4 in). Cetakan-cetakan tersebut berantakan, dan lebarnya cetakan jari kaki yang bervariasi mengindikasikan bahwa lumpur telah menempel pada kakinya. Bentuk cetakannya bervariasi menurut perbedaan pada substrat dan bagaimana cetakan tersebut dibuat; terkadang, kakinya dijejakkan secara langsung, tetapi sering kali terjadi selipan ke belakang maupun ke depan saat kakinya turun berpijak. Posisi dan sudut jari-jarinya juga sangat bervariasi, yang mengindikasikan bahwa jari-jari itu pastilah cukup fleksibel. Jejak kaki Dilophosauripus memiliki jari kaki kedua yang tergeser dengan pangkal yang tebal, dan cakar yang sangat panjang serta lurus yang sejajar dengan poros bantalan jarinya. Salah satu jejak kakinya kehilangan cakar di jari kedua, yang mungkin diakibatkan oleh cedera.[40] Pada tahun 1984, Welles menginterpretasikan fakta bahwa tiga individu ditemukan saling berdekatan, serta adanya lintasan jejak yang saling menyilang di dekatnya, sebagai indikasi bahwa Dilophosaurus bepergian secara berkelompok.[2] Gay sepakat bahwa mereka mungkin bepergian dalam kelompok-kelompok kecil, tetapi ia mencatat bahwa belum ada bukti langsung yang mendukung hal ini, dan bahwa banjir bandang dapat saja menyapu tulang-belulang yang berserakan dari individu-individu yang berbeda dan mengendapkannya secara bersama-sama.[12]
Milner dan rekan-rekannya memeriksa kemungkinan lintasan jejak Dilophosaurus SGDS 18.T1 pada tahun 2009, yang terdiri dari jejak kaki khas dengan seretan ekor dan jejak beristirahat yang lebih tidak biasa, yang terendapkan di batu pasir pantai lakustrin (danau). Lintasan tersebut dimulai dengan hewan yang awalnya menghadap secara kasarnya sejajar dengan garis pantai, dan kemudian berhenti di sebuah berm (tanggul) dengan kedua kakinya sejajar, yang setelah itu ia menurunkan badannya, dan menempatkan metatarsal serta kalus di sekitar iskiumnya ke tanah; hal ini menciptakan cetakan "tumit" yang simetris dan cetakan melingkar dari iskium. Bagian ekor yang terdekat dengan badan ditahan agar tidak menyentuh tanah, sedangkan bagian ujung yang lebih jauh dari badan bersentuhan dengan tanah. Fakta bahwa hewan tersebut beristirahat di suatu lereng memungkinkannya meletakkan kedua tangannya ke tanah di dekat kakinya. Setelah beristirahat, dinosaurus itu menyeret kakinya ke depan, dan meninggalkan cetakan baru menggunakan kaki, metatarsal, dan iskiumnya, tanpa mengikutsertakan tangannya. Kaki kanannya kini menginjak cetakan tangan kanannya, dan cakar kedua dari kaki kirinya membuat bekas seretan dari posisi istirahat pertama ke posisi istirahat selanjutnya. Setelah beberapa saat, hewan tersebut berdiri dan bergerak maju, didahului dengan kaki kirinya, dan setelah berdiri tegak sepenuhnya, ia berjalan melintasi sisa permukaan tanah yang terbuka, sembari meninggalkan bekas seretan tipis dengan ujung ekornya.[49]
Meringkuk merupakan perilaku teropoda yang jarang tertangkap, dan SGDS 18.T1 adalah satu-satunya jejak sedemikian rupa dengan cetakan tanpa ambigu dari tangan teropoda, yang memberikan informasi berharga mengenai bagaimana hewan ini menggunakan tungkai depannya. Postur meringkuknya didapati sangat mirip dengan postur burung modern, dan menunjukkan bahwa teropoda awal menahan telapak tangan mereka agar berhadapan ke arah medial, saling berhadapan satu sama lain. Karena postur semacam ini berevolusi pada tahap awal garis keturunannya, hal itu mungkin telah menjadi ciri khas bagi seluruh teropoda. Teropoda sering kali digambarkan dengan telapak tangan yang menghadap ke bawah, namun penelitian atas anatomi fungsionalnya telah membuktikan bahwa mereka, layaknya burung, tidak dapat mempronasi maupun menyupinasi lengannya. Jejak tersebut menunjukkan bahwa kakinya ditahan secara simetris dengan bobot tubuh yang terdistribusikan di antara kaki dan metatarsal, yang mana ini juga merupakan ciri yang terlihat pada burung seperti ratit. Milner dan rekan-rekannya juga menepis gagasan bahwa jejak Kayentapus minor yang dilaporkan oleh Weems menunjukkan cetakan telapak tangan yang dibuat oleh teropoda yang berjalan secara kuadrupedal. Weems telah mengusulkan pembuat jejak itu mungkin mampu bergerak secara kuadrupedal saat berjalan pelan, sementara jari-jarinya akan selalu secara terbiasa dihiperekstensikan sehingga hanya telapak tangannya yang menyentuh tanah. Milner dan rekan-rekannya menganggap bahwa penyimpulan pose ini tidaklah perlu, dan menyarankan bahwa jejak tersebut sebaliknya dibuat dengan cara yang serupa seperti pada SGDS 18.T1, namun tanpa meninggalkan bekas jari kaki.[49]
Fungsi jambul
sunting
Welles mengakui bahwa usulan mengenai fungsi jambul Dilophosaurus bersifat konjektural (dugaan), tetapi ia berpendapat bahwa, meskipun jambul tersebut tidak memiliki alur yang mengindikasikan vaskularisasi, struktur tersebut bisa saja digunakan untuk termoregulasi. Ia juga menyarankan bahwa jambul itu mungkin digunakan untuk pengenalan spesies atau ornamentasi.[2] Bakker menganggap jambul tersebut sebagai perhiasan seksual pada tahun 1986, dengan menyoroti bahwa jambul itu sangat tipis sehingga hanya mungkin berfungsi untuk efek visual, berbeda dengan jambul alosaurus yang lebih berat, yang bisa jadi digunakan untuk saling membenturkan kepala.[52] Czerkas bersaudara menunjukkan pada tahun 1990 bahwa jambul tersebut tidak mungkin digunakan dalam pertarungan, karena strukturnya yang rapuh akan mudah rusak. Mereka mengusulkan bahwa jambul itu merupakan pertunjukan visual untuk menarik pasangan, dan bahkan termoregulasi.[56] Pada tahun 1990, ahli paleontologi Walter P. Coombs menyatakan bahwa jambul tersebut mungkin dipertegas oleh warna untuk digunakan dalam pertunjukan pamer.[62]
Pada tahun 2011, ahli paleontologi Kevin Padian dan John R. Horner mengusulkan bahwa "struktur aneh" pada dinosaurus secara umum (termasuk jambul, jumbai, tanduk, dan kubah) utamanya digunakan untuk pengenalan spesies, dan menepis penjelasan lain karena tidak didukung oleh bukti. Mereka menyoroti bahwa terlalu sedikit spesimen teropoda berornamen kranial, termasuk Dilophosaurus, yang diketahui untuk dapat menguji fungsi evolusionernya secara statistik, dan apakah struktur tersebut merepresentasikan dimorfisme seksual atau kematangan seksual.[63] Sebagai tanggapan terhadap Padian dan Horner pada tahun yang sama, ahli paleontologi Rob J. Knell dan Scott D. Sampson berargumen bahwa pengenalan spesies bukanlah hal yang tidak mungkin sebagai fungsi sekunder untuk "struktur aneh" pada dinosaurus, melainkan bahwa seleksi seksual (digunakan dalam pertunjukan pamer atau pertarungan untuk memperebutkan pasangan) merupakan penjelasan yang lebih masuk akal, mengingat tingginya biaya biologis untuk mengembangkannya, dan karena struktur semacam itu tampaknya sangat bervariasi di dalam suatu spesies.[64]
Pada tahun 2013, ahli paleontologi David E. Hone dan Darren Naish mengkritik "hipotesis pengenalan spesies", dan berargumen bahwa tidak ada hewan yang masih ada saat ini menggunakan struktur semacam itu utamanya untuk pengenalan spesies, dan bahwa Padian dan Horner telah mengabaikan kemungkinan seleksi seksual timbal balik (di mana kedua jenis kelamin memiliki ornamen).[65] Marsh dan Rowe sepakat pada tahun 2020 bahwa jambul Dilophosaurus kemungkinan memiliki peran dalam identifikasi spesies atau seleksi interseksual/intraseksual, seperti pada beberapa burung modern.[6] Tidak diketahui apakah kantung udara di dalam jambul mendukung fungsi-fungsi tersebut.[15]
Perkembangan
sunting
Welles pada awalnya menginterpretasikan spesimen Dilophosaurus yang lebih kecil sebagai individu remaja, dan spesimen yang lebih besar sebagai individu dewasa, namun kemudian menginterpretasikan mereka sebagai spesies yang berbeda.[2][7] Paul menyarankan bahwa perbedaan di antara spesimen-spesimen tersebut mungkin disebabkan oleh dimorfisme seksual, sebagaimana yang tampaknya juga terlihat pada Coelophysis, yang memiliki bentuk "tegap" dan "ramping" dengan ukuran yang sama, yang jika tidak demikian mungkin akan dianggap sebagai spesies terpisah. Mengikuti skema ini, spesimen Dilophosaurus yang lebih kecil akan merepresentasikan contoh yang "ramping".[24]
Pada tahun 2005 Tykoski menemukan bahwa sebagian besar spesimen Dilophosaurus yang diketahui adalah individu remaja, dengan hanya spesimen terbesar yang merupakan individu dewasa, berdasarkan tingkat ko-osifikasi tulang-tulangnya.[14] Pada tahun 2005 Gay tidak menemukan bukti adanya dimorfisme seksual seperti yang diusulkan oleh Paul (namun diduga hadir pada Coelophysis), dan mengaitkan variasi yang terlihat di antara spesimen Dilophosaurus dengan variasi individual dan ontogeni (perubahan selama masa pertumbuhan). Tidak ada dimorfisme pada kerangkanya, tetapi ia tidak menyingkirkan kemungkinan bahwa hal itu bisa saja terdapat pada jambulnya; diperlukan lebih banyak data untuk menentukannya.[17] Berdasarkan jambul hidung berukuran kecil pada sebuah spesimen remaja, yang sebelumnya secara tentatif dimasukkan oleh Yates ke dalam genus kerabat Dracovenator, ia mengusulkan bahwa jambul ini akan tumbuh lebih besar seiring hewan tersebut menjadi dewasa.[36]
Ahli paleontologi J.S. Tkach melaporkan sebuah studi histologis (studi mikroskopis terhadap ciri internal) dari Dilophosaurus pada tahun 1996, yang dilakukan dengan mengambil sayatan tipis dari tulang panjang dan tulang rusuk spesimen UCMP 37303 (kerangka yang paling tidak terawetkan di antara dua kerangka aslinya). Jaringan tulangnya tervaskularisasi dengan baik dan memiliki struktur fibro-lamelar yang mirip dengan yang ditemukan pada teropoda lain dan sauropodomorfa Massospondylus. Struktur pleksiform (anyaman) pada tulangnya mengindikasikan pertumbuhan yang cepat, dan Dilophosaurus mungkin mencapai tingkat pertumbuhan 30 hingga 35 kilogram (66 hingga 77 pon) per tahun pada awal kehidupannya.[66]
Welles menemukan bahwa gigi pengganti Dilophosaurus dan teropoda lain bermula dari dalam tulang, dengan ukuran yang menyusut semakin jauh jaraknya dari batas alveolar. Biasanya terdapat dua atau tiga gigi pengganti di dalam alveoli, dengan gigi termuda berupa mahkota gigi yang kecil dan berongga. Gigi pengganti erupsi di sisi luar gigi yang lama. Ketika sebuah gigi mendekati garis gusi, dinding bagian dalam di antara lempeng interdental diserap kembali dan membentuk takik nutrien. Seiring erupsi gigi yang baru, gigi tersebut bergerak ke luar untuk memosisikan dirinya di tengah alveolus, dan takik nutrien tersebut pun menutup.[2]
Paleopatologi
sunting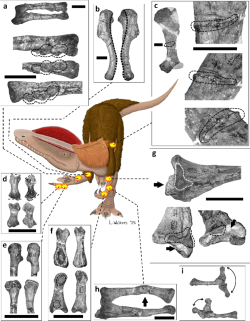
Welles mencatat berbagai paleopatologi (tanda-tanda kuno adanya penyakit, seperti cedera dan malformasi) pada Dilophosaurus. Spesimen holotipe memiliki sebuah sulkus (alur atau lekukan) pada lengkungan saraf dari tulang belakang servikal yang mungkin disebabkan oleh cedera atau remukan, serta dua lubang pada humerus kanan yang mungkin merupakan abses (kumpulan nanah) atau artefak. Welles juga mencatat bahwa hewan ini memiliki humerus kiri yang lebih kecil dan tipis daripada yang kanan, namun dengan kondisi yang berkebalikan pada lengan bawahnya. Pada tahun 2001, ahli paleontologi Ralph Molnar menyarankan bahwa hal ini disebabkan oleh anomali perkembangan yang disebut asimetri fluktuatif. Anomali ini dapat disebabkan oleh stres pada populasi hewan, misalnya akibat gangguan di lingkungan mereka, dan mungkin mengindikasikan tekanan selektif yang lebih intens. Asimetri juga dapat diakibatkan oleh peristiwa traumatis pada awal masa perkembangan seekor hewan, yang mana akan terdistribusi secara lebih acak dalam hal waktu.[2][67] Sebuah studi tahun 2001 yang dilakukan oleh ahli paleontologi Bruce Rothschild dan rekan-rekannya memeriksa 60 tulang kaki Dilophosaurus untuk mencari tanda-tanda fraktur stres (yang disebabkan oleh tindakan berat dan berulang), tetapi tidak ditemukan satu pun. Cedera semacam itu dapat merupakan akibat dari gaya hidup predator yang sangat aktif.[68]
Pada tahun 2016 Senter dan Sara L. Juengst memeriksa paleopatologi dari spesimen holotipe dan menemukan bahwa spesimen tersebut menanggung jumlah penyakit terbanyak dan paling beragam di gelang bahu dan tungkai depannya dibandingkan dengan dinosaurus teropoda mana pun yang telah dideskripsikan sejauh ini, yang beberapa di antaranya tidak diketahui dari dinosaurus lainnya. Hanya enam teropoda lain yang diketahui memiliki lebih dari satu paleopatologi di gelang bahu dan tungkai depan. Spesimen holotipe memiliki delapan tulang yang terserang penyakit, sedangkan tidak ada spesimen teropoda lain yang diketahui memiliki lebih dari empat. Di sisi kirinya, ia mengalami patah tulang skapula dan radius, serta fibrisces (sejenis abses) di ulna dan tulang falang terluar pada ibu jari. Di sisi kanannya, ia mengalami torsi (pelintiran) pada batang humerusnya, tiga tumor tulang pada radiusnya, permukaan artikular yang terpotong pada tulang metakarpal ketiganya, dan kelainan bentuk pada tulang falang pertama di jari ketiganya. Jari ketiga ini cacat permanen dan tidak dapat ditekuk. Kelainan bentuk pada humerus dan jari ketiga mungkin disebabkan oleh osteodisplasia, yang sebelumnya belum pernah dilaporkan pada dinosaurus non-burung, tetapi lazim diketahui pada burung. Menjangkiti burung remaja yang mengalami kekurangan gizi, penyakit ini dapat menyebabkan rasa sakit pada salah satu tungkai, yang membuat burung lebih memilih untuk menggunakan tungkai lainnya, yang karenanya tungkai tersebut mengalami torsi.[69]

Jumlah peristiwa traumatis yang menyebabkan ciri-ciri ini tidaklah pasti, dan mungkin saja semuanya disebabkan oleh satu kejadian tunggal, misalnya akibat menabrak pohon atau batu selama pertarungan dengan hewan lain, yang mungkin telah menyebabkan luka tusuk dari cakarnya. Karena semua cederanya telah sembuh, sudah dapat dipastikan bahwa Dilophosaurus tersebut sintas dalam waktu yang lama setelah peristiwa-peristiwa tersebut, selama berbulan-bulan, mungkin bertahun-tahun. Penggunaan tungkai depan untuk menangkap mangsa pastilah terganggu selama masa penyembuhan. Oleh karena itu, dinosaurus tersebut mungkin mengalami masa puasa yang panjang atau bertahan hidup dengan memangsa hewan yang cukup kecil untuk dihabisi menggunakan mulut dan kakinya, atau menggunakan salah satu tungkai depannya.[69]
Menurut Senter dan Juengst, tingkat rasa sakit yang tinggi yang mungkin dialami dinosaurus tersebut di beberapa lokasi tubuh selama durasi yang panjang juga menunjukkan bahwa ia adalah hewan yang tangguh. Mereka mencatat bahwa paleopatologi pada dinosaurus kurang banyak dilaporkan, dan meskipun Welles telah mendeskripsikan spesimen holotipe secara menyeluruh, ia hanya menyebutkan satu dari patologi-patologi yang mereka temukan. Mereka menyarankan bahwa ciri-ciri semacam itu terkadang mungkin dihilangkan karena deskripsi spesies lebih berfokus pada karakteristik mereka ketimbang keabnormalannya, atau karena ciri-ciri tersebut sulit untuk dikenali.[69] Senter dan Sullivan menemukan bahwa patologi-patologi tersebut secara signifikan mengubah rentang gerak di bahu kanan dan jari ketiga sebelah kanan pada spesimen holotipe, dan bahwa perkiraan rentang gerak tersebut oleh karenanya mungkin tidak sama dengan apa yang diukur pada tungkai depan yang sehat.[61]
Perkembangan
suntingWelles pada awalnya menginterpretasikan spesimen Dilophosaurus yang lebih kecil sebagai individu remaja, dan spesimen yang lebih besar sebagai individu dewasa, namun kemudian menginterpretasikan mereka sebagai spesies yang berbeda.[2][7] Paul menyarankan bahwa perbedaan di antara spesimen-spesimen tersebut mungkin disebabkan oleh dimorfisme seksual, sebagaimana yang tampaknya juga terlihat pada Coelophysis, yang memiliki bentuk "tegap" dan "ramping" dengan ukuran yang sama, yang jika tidak demikian mungkin akan dianggap sebagai spesies terpisah. Mengikuti skema ini, spesimen Dilophosaurus yang lebih kecil akan merepresentasikan contoh yang "ramping".[24]
Pada tahun 2005 Tykoski menemukan bahwa sebagian besar spesimen Dilophosaurus yang diketahui adalah individu remaja, dengan hanya spesimen terbesar yang merupakan individu dewasa, berdasarkan tingkat ko-osifikasi tulang-tulangnya.[14] Pada tahun 2005 Gay tidak menemukan bukti adanya dimorfisme seksual seperti yang diusulkan oleh Paul (namun diduga hadir pada Coelophysis), dan mengaitkan variasi yang terlihat di antara spesimen Dilophosaurus dengan variasi individual dan ontogeni (perubahan selama masa pertumbuhan). Tidak ada dimorfisme pada kerangkanya, tetapi ia tidak menyingkirkan kemungkinan bahwa hal itu bisa saja terdapat pada jambulnya; diperlukan lebih banyak data untuk menentukannya.[17] Berdasarkan jambul hidung berukuran kecil pada sebuah spesimen remaja, yang sebelumnya secara tentatif dimasukkan oleh Yates ke dalam genus kerabat Dracovenator, ia mengusulkan bahwa jambul ini akan tumbuh lebih besar seiring hewan tersebut menjadi dewasa.[36]
Ahli paleontologi J.S. Tkach melaporkan sebuah studi histologis (studi mikroskopis terhadap ciri internal) dari Dilophosaurus pada tahun 1996, yang dilakukan dengan mengambil sayatan tipis dari tulang panjang dan tulang rusuk spesimen UCMP 37303 (kerangka yang paling tidak terawetkan di antara dua kerangka aslinya). Jaringan tulangnya tervaskularisasi dengan baik dan memiliki struktur fibro-lamelar yang mirip dengan yang ditemukan pada teropoda lain dan sauropodomorfa Massospondylus. Struktur pleksiform (anyaman) pada tulangnya mengindikasikan pertumbuhan yang cepat, dan Dilophosaurus mungkin mencapai tingkat pertumbuhan 30 hingga 35 kilogram (66 hingga 77 pon) per tahun pada awal kehidupannya.[66]
Welles menemukan bahwa gigi pengganti Dilophosaurus dan teropoda lain bermula dari dalam tulang, dengan ukuran yang menyusut semakin jauh jaraknya dari batas alveolar. Biasanya terdapat dua atau tiga gigi pengganti di dalam alveoli, dengan gigi termuda berupa mahkota gigi yang kecil dan berongga. Gigi pengganti erupsi di sisi luar gigi yang lama. Ketika sebuah gigi mendekati garis gusi, dinding bagian dalam di antara lempeng interdental diserap kembali dan membentuk takik nutrien. Seiring erupsi gigi yang baru, gigi tersebut bergerak ke luar untuk memosisikan dirinya di tengah alveolus, dan takik nutrien tersebut pun menutup.[2]
Paleopatologi
suntingWelles mencatat berbagai paleopatologi (tanda-tanda kuno adanya penyakit, seperti cedera dan malformasi) pada Dilophosaurus. Spesimen holotipe memiliki sebuah sulkus (alur atau lekukan) pada lengkungan saraf dari tulang belakang servikal yang mungkin disebabkan oleh cedera atau remukan, serta dua lubang pada humerus kanan yang mungkin merupakan abses (kumpulan nanah) atau artefak. Welles juga mencatat bahwa hewan ini memiliki humerus kiri yang lebih kecil dan tipis daripada yang kanan, namun dengan kondisi yang berkebalikan pada lengan bawahnya. Pada tahun 2001, ahli paleontologi Ralph Molnar menyarankan bahwa hal ini disebabkan oleh anomali perkembangan yang disebut asimetri fluktuatif. Anomali ini dapat disebabkan oleh stres pada populasi hewan, misalnya akibat gangguan di lingkungan mereka, dan mungkin mengindikasikan tekanan selektif yang lebih intens. Asimetri juga dapat diakibatkan oleh peristiwa traumatis pada awal masa perkembangan seekor hewan, yang mana akan terdistribusi secara lebih acak dalam hal waktu.[2][67] Sebuah studi tahun 2001 yang dilakukan oleh ahli paleontologi Bruce Rothschild dan rekan-rekannya memeriksa 60 tulang kaki Dilophosaurus untuk mencari tanda-tanda fraktur stres (yang disebabkan oleh tindakan berat dan berulang), tetapi tidak ditemukan satu pun. Cedera semacam itu dapat merupakan akibat dari gaya hidup predator yang sangat aktif.[68]
Pada tahun 2016 Senter dan Sara L. Juengst memeriksa paleopatologi dari spesimen holotipe dan menemukan bahwa spesimen tersebut menanggung jumlah penyakit terbanyak dan paling beragam di gelang bahu dan tungkai depannya dibandingkan dengan dinosaurus teropoda mana pun yang telah dideskripsikan sejauh ini, yang beberapa di antaranya tidak diketahui dari dinosaurus lainnya. Hanya enam teropoda lain yang diketahui memiliki lebih dari satu paleopatologi di gelang bahu dan tungkai depan. Spesimen holotipe memiliki delapan tulang yang terserang penyakit, sedangkan tidak ada spesimen teropoda lain yang diketahui memiliki lebih dari empat. Di sisi kirinya, ia mengalami patah tulang skapula dan radius, serta fibrisces (sejenis abses) di ulna dan tulang falang terluar pada ibu jari. Di sisi kanannya, ia mengalami torsi (pelintiran) pada batang humerusnya, tiga tumor tulang pada radiusnya, permukaan artikular yang terpotong pada tulang metakarpal ketiganya, dan kelainan bentuk pada tulang falang pertama di jari ketiganya. Jari ketiga ini cacat permanen dan tidak dapat ditekuk. Kelainan bentuk pada humerus dan jari ketiga mungkin disebabkan oleh osteodisplasia, yang sebelumnya belum pernah dilaporkan pada dinosaurus non-burung, tetapi lazim diketahui pada burung. Menjangkiti burung remaja yang mengalami kekurangan gizi, penyakit ini dapat menyebabkan rasa sakit pada salah satu tungkai, yang membuat burung lebih memilih untuk menggunakan tungkai lainnya, yang karenanya tungkai tersebut mengalami torsi.[69]
Jumlah peristiwa traumatis yang menyebabkan ciri-ciri ini tidaklah pasti, dan mungkin saja semuanya disebabkan oleh satu kejadian tunggal, misalnya akibat menabrak pohon atau batu selama pertarungan dengan hewan lain, yang mungkin telah menyebabkan luka tusuk dari cakarnya. Karena semua cederanya telah sembuh, sudah dapat dipastikan bahwa Dilophosaurus tersebut sintas dalam waktu yang lama setelah peristiwa-peristiwa tersebut, selama berbulan-bulan, mungkin bertahun-tahun. Penggunaan tungkai depan untuk menangkap mangsa pastilah terganggu selama masa penyembuhan. Oleh karena itu, dinosaurus tersebut mungkin mengalami masa puasa yang panjang atau bertahan hidup dengan memangsa hewan yang cukup kecil untuk dihabisi menggunakan mulut dan kakinya, atau menggunakan salah satu tungkai depannya.[69]
Menurut Senter dan Juengst, tingkat rasa sakit yang tinggi yang mungkin dialami dinosaurus tersebut di beberapa lokasi tubuh selama durasi yang panjang juga menunjukkan bahwa ia adalah hewan yang tangguh. Mereka mencatat bahwa paleopatologi pada dinosaurus kurang banyak dilaporkan, dan meskipun Welles telah mendeskripsikan spesimen holotipe secara menyeluruh, ia hanya menyebutkan satu dari patologi-patologi yang mereka temukan. Mereka menyarankan bahwa ciri-ciri semacam itu terkadang mungkin dihilangkan karena deskripsi spesies lebih berfokus pada karakteristik mereka ketimbang keabnormalannya, atau karena ciri-ciri tersebut sulit untuk dikenali.[69] Senter dan Sullivan menemukan bahwa patologi-patologi tersebut secara signifikan mengubah rentang gerak di bahu kanan dan jari ketiga sebelah kanan pada spesimen holotipe, dan bahwa perkiraan rentang gerak tersebut oleh karenanya mungkin tidak sama dengan apa yang diukur pada tungkai depan yang sehat.[61]
Lingkungan purba
sunting
Dilophosaurus diketahui dari Formasi Kayenta, yang berasal dari tahap Sinemurium-Toarkium pada masa Jura Awal, sekitar 196–186 juta tahun yang lalu (187–190 juta tahun yang lalu juga telah diusulkan, dan umur Formasi Kayenta dianggap kompleks). Karena Dilophosaurus diketahui dari bagian dasar hingga bagian tengah formasi, yang berumur Pliensbakium, takson ini memiliki rentang kronostratigrafis selama 15 juta tahun.[70][71] Formasi Kayenta adalah bagian dari Kelompok Glen Canyon yang mencakup formasi-formasi di Arizona utara, sebagian tenggara Utah, Colorado barat, dan New Mexico barat laut. Formasi ini sebagian besar tersusun atas dua fasies, yang satu didominasi oleh endapan batu lanau dan yang lainnya oleh batu pasir. Fasies batu lanau banyak ditemukan di Arizona, sedangkan fasies batu pasir hadir di wilayah Arizona utara, Utah selatan, Colorado barat, dan New Mexico barat laut. Formasi ini utamanya diendapkan oleh sungai-sungai, dengan fasies batu lanau sebagai bagian yang lebih lambat dan tenang dari sistem sungai tersebut. Pengendapan Formasi Kayenta diakhiri oleh perambahan padang gumuk yang kelak menjadi Batu Pasir Navajo.[72] Lingkungan tersebut mengalami musim kering musiman, dengan gumuk pasir yang bermigrasi masuk dan keluar dari lingkungan basah tempat hewan-hewan hidup, dan telah diibaratkan sebagai oasis sungai; sebuah jalur air yang ditumbuhi deretan tumbuhan runjung dan dikelilingi oleh pasir. [15]
Formasi Kayenta telah menghasilkan kumpulan organisme yang kecil namun terus bertambah. Sebagian besar fosil berasal dari fasies batu lanau.[73] Sebagian besar organisme yang diketahui sejauh ini adalah vertebrata. Invertebrata dan organisme non-vertebrata lainnya mencakup batu kapur mikrob atau "alga",[74] kayu yang membatu,[75] cetakan tumbuhan,[76] kerang (bivalvia) dan siput air tawar,[72] ostrakoda,[77] serta fosil jejak invertebrata.[74] Vertebrata diketahui baik dari fosil tubuh maupun fosil jejak. Vertebrata yang diketahui dari fosil tubuh mencakup[73] hiu hibodon, ikan bertulang sejati (osteichthyes) yang belum teridentifikasi, ikan paru-paru,[75] salamander,[78] katak Prosalirus, sesilia Eocaecilia, kura-kura Kayentachelys, reptil sfenodon, kadal,[79] dan beberapa krokodilomorf awal termasuk Calsoyasuchus, Eopneumatosuchus, Kayentasuchus, dan Protosuchus, serta pterosaurus Rhamphinion. Selain Dilophosaurus, beberapa dinosaurus juga diketahui, termasuk teropoda Megapnosaurus,[14] dan Kayentavenator,[80] sauropodomorfa Sarahsaurus,[81] seekor heterodontosaurid, dan tireoforan Scutellosaurus. Sinapsida mencakup tritilodontid Dinnebitodon, Kayentatherium, dan Oligokyphus, morganukodontid,[79] kemungkinan mamalia sejati awal Dinnetherium, dan mamalia haramiid. Mayoritas temuan ini berasal dari sekitar Gold Spring, Arizona.[73] Fosil jejak vertebrata mencakup koprolit dan jejak kaki terapsida, hewan mirip kadal, serta beberapa jenis dinosaurus.[74][82]
Tafonomi
suntingWelles menguraikan tafonomi dari spesimen aslinya, yaitu perubahan-perubahan yang terjadi selama pembusukan dan fosilisasinya. Kerangka holotipe ditemukan berbaring miring di sisi kanannya, dan kepala serta lehernya melengkung ke belakang – tertarik ke belakang – dalam "pose kematian" yang mana kerangka dinosaurus sering kali ditemukan dalam pose tersebut. Pose ini pada saat itu diduga sebagai opistotonus (akibat kejang menjelang kematian), namun mungkin saja merupakan akibat dari bagaimana bangkai tersebut tertanam di dalam sedimen. Punggungnya lurus, dan tulang belakang dorsal (punggung) paling belakang memutar ke sisi kirinya. Tulang belakang kaudal (ekor) memanjang secara tak beraturan dari panggul, dan kaki-kakinya terartikulasi, dengan sedikit pergeseran letak. Welles menyimpulkan bahwa spesimen-spesimen tersebut terkubur di tempat kematian mereka, tanpa banyak berpindah tempat (tertransportasi), tetapi spesimen holotipe tampaknya telah diganggu oleh pemakan bangkai, yang diindikasikan oleh rotasi pada tulang belakang dorsal dan tengkoraknya yang hancur.[2][83] Gay mencatat bahwa spesimen-spesimen yang ia deskripsikan pada tahun 2001 menunjukkan bukti pernah tertransportasi oleh aliran sungai. Karena tak satupun dari spesimen tersebut yang lengkap, mereka mungkin telah tertransportasi dalam jarak tertentu, atau sempat tergeletak di permukaan tanah dan melapuk untuk beberapa waktu sebelum akhirnya tertransportasi. Mereka mungkin telah terbawa oleh banjir, yang diindikasikan oleh keberagaman hewan yang ditemukan dalam bentuk fragmen dan pecahan tulang.[12]
Signifikansi budaya
sunting

Menurut mitologi Navajo, bangkai monster yang terbunuh "dibenamkan ke dalam bumi", tetapi mustahil untuk dilenyapkan, dan fosil secara tradisional telah diinterpretasikan sebagai sisa-sisanya. Meskipun masyarakat Navajo telah membantu para ahli paleontologi menemukan lokasi fosil sejak abad ke-19, kepercayaan tradisional menunjukkan bahwa hantu dari monster-monster tersebut tetap berada di dalam mayat mereka yang setengah terkubur, dan harus ditahan di sana melalui ritual-ritual yang kuat. Demikian pula, beberapa orang khawatir bahwa tulang-belulang kerabat mereka akan ikut tergali bersama dengan sisa-sisa dinosaurus, dan bahwa memindahkan fosil menunjukkan rasa tidak hormat terhadap kehidupan masa lalu dari makhluk-makhluk ini.[84] Pada tahun 2005, sejarawan Adrienne Mayor menyatakan bahwa Welles telah mencatat bahwa selama penggalian asli Dilophosaurus, orang Navajo bernama Williams menghilang dari penggalian setelah beberapa hari, dan berspekulasi bahwa hal ini terjadi karena Williams menganggap pekerjaan mendetail menggunakan kuas halus "merendahkan martabatnya". Mayor sebaliknya menunjukkan bahwa para pria Navajo memang menyibukkan diri dengan pekerjaan mendetail, seperti pembuatan perhiasan dan lukisan, dan bahwa penjelasan atas kepergian Williams mungkin sebaliknya adalah kecemasan tradisional saat kerangka tersebut muncul ke permukaan dan diganggu. Mayor juga menunjuk pada sebuah insiden pada tahun 1940-an ketika seorang pria Navajo membantu menggali kerangka Pentaceratops selama ia tidak harus menyentuh tulang-tulangnya, namun meninggalkan situs tersebut ketika hanya tinggal beberapa inci tanah yang menutupinya.[84] Dalam sebuah buku tahun 1994, Welles mengatakan bahwa Williams telah kembali beberapa hari kemudian bersama dua wanita Navajo dan mengatakan "itu bukan pekerjaan pria, itu pekerjaan wanita (squaw)".[9]
Tebing-tebing di Arizona yang menyimpan tulang-belulang Dilophosaurus juga memiliki petroglif buatan orang Pueblo leluhur yang terukir di atasnya, dan jejak-jejak yang saling menyilang di area tersebut disebut Naasho'illbahitsho Biikee oleh orang Navajo, yang berarti "jejak kaki kadal besar". Menurut Mayor, orang Navajo dulunya mengadakan upacara dan memberikan persembahan pada jejak-jejak monster ini. Jejak kaki tridaktil juga ditampilkan sebagai dekorasi pada pakaian dan seni cadas dari masyarakat Hopi dan Zuni, yang kemungkinan terpengaruh oleh jejak dinosaurus semacam itu.[84] Pada tahun 2017 Dilophosaurus ditetapkan sebagai dinosaurus negara bagian di negara bagian AS Connecticut, yang menjadi resmi melalui anggaran negara bagian baru pada tahun 2019. Dilophosaurus dipilih karena jejak kaki yang diduga dibuat oleh dinosaurus serupa ditemukan di Rocky Hill pada tahun 1966, selama penggalian untuk Interstate Highway 91. Keenam jejak tersebut dikaitkan ke dalam iknospesies Eubrontes giganteus, yang dijadikan fosil negara bagian Connecticut pada tahun 1991. Area tempat jejak-jejak itu ditemukan dulunya merupakan danau masa Trias, dan ketika signifikansi area tersebut dikonfirmasi, jalan raya itu dialihkan rutenya, dan area tersebut dijadikan taman negara bagian yang diberi nama Dinosaur State Park. Pada tahun 1981, sebuah patung Dilophosaurus, yakni rekonstruksi seukuran aslinya yang pertama untuk dinosaurus ini, didonasikan ke taman tersebut.[24][85][86]
Dilophosaurus diusulkan sebagai dinosaurus negara bagian Arizona oleh seorang anak laki-laki berusia sembilan tahun pada tahun 1998, namun para pembuat undang-undang menyarankan Sonorasaurus sebagai gantinya, dengan argumen bahwa Dilophosaurus tidak unik bagi Arizona. Sebuah kompromi sempat disarankan untuk mengakui kedua dinosaurus tersebut, namun rancangan undang-undang itu dibatalkan ketika terungkap bahwa fosil Dilophosaurus tersebut telah diambil tanpa izin dari Reservasi Navajo, dan karena fosil-fosil tersebut tidak lagi berada di Arizona (seorang anak laki-laki berusia 11 tahun kembali mengusulkan Sonorasaurus sebagai dinosaurus negara bagian Arizona pada tahun 2018). Para pejabat Bangsa Navajo kemudian mendiskusikan cara agar fosil-fosil tersebut dikembalikan.[87][88] Menurut Mayor, seorang Navajo menyatakan bahwa mereka tidak lagi meminta fosil tersebut dikembalikan, tetapi mempertanyakan mengapa tidak dibuatkan cetakannya saja agar tulang-tulang aslinya dapat ditinggalkan, karena akan lebih baik menyimpannya di dalam tanah, dan sebuah museum dibangun agar orang-orang dapat datang untuk melihatnya di sana.[84] Kerja lapangan lebih lanjut yang berkaitan dengan Dilophosaurus di Bangsa Navajo dilakukan dengan izin dari Departemen Mineral Bangsa Navajo.[15]
Jurassic Park
sunting
Dilophosaurus ditampilkan dalam novel tahun 1990 Jurassic Park, oleh penulis Michael Crichton, dan adaptasi filmnya tahun 1993 oleh sutradara Steven Spielberg. Dilophosaurus di Jurassic Park diakui sebagai "satu-satunya penyimpangan serius dari kebenaran ilmiah" di dalam buku pembuatan film tersebut, dan sebagai dinosaurus "yang paling difiksikan" di film tersebut dalam sebuah buku tentang Stan Winston Studios, yang menciptakan efek animatroniknya. Untuk novelnya, Crichton menciptakan kemampuan dinosaurus tersebut untuk meludahkan bisa (yang menjelaskan bagaimana ia mampu membunuh mangsa, terlepas dari rahangnya yang terlihat lemah). Departemen seninya menambahkan fitur lain, yaitu jumbai leher atau kerudung yang terlipat di lehernya yang mengembang dan bergetar saat hewan itu bersiap untuk menyerang, mirip dengan yang dimiliki oleh kadal berjumbai. Untuk menghindari kebingungan dengan Velociraptor yang ditampilkan di dalam film, Dilophosaurus disajikan dengan tinggi yang hanya 1,2 meter (4 ft), bukan dari tinggi asli yang diasumsikan yakni sekitar 3,0 meter (10 ft). Dijuluki "si peludah", Dilophosaurus dalam film ini direalisasikan melalui teknik pewayangan (puppeteering), dan membutuhkan satu tubuh penuh dengan tiga kepala yang dapat diganti-ganti untuk menghasilkan aksi yang diperlukan oleh naskah. Kepala pertama memiliki jumbai yang terlipat, kepala kedua dapat mengembangkan dan melipat jumbainya, dan kepala ketiga memiliki jumbai yang terentang dan bergetar serta dapat meludahkan bisa. Kaki yang terpisah juga dibuat untuk adegan di mana dinosaurus tersebut melompat lewat. Tidak seperti sebagian besar dinosaurus lain dalam film tersebut, tidak ada citra hasil komputer (CGI) yang digunakan saat menampilkan Dilophosaurus.[89][90][91]
Ahli geologi J. Bret Bennington mencatat pada tahun 1996 bahwa meskipun Dilophosaurus kemungkinan besar tidak memiliki jumbai dan tidak dapat meludahkan bisa seperti di dalam film, gigitannya bisa saja berbisa, seperti yang diklaim ada pada biawak komodo. Ia berpendapat bahwa menambahkan fitur bisa pada dinosaurus ini sama diizinkannya dengan memberikan warna pada kulitnya, yang mana juga tidak diketahui. Jika dinosaurus ini memiliki jumbai, pasti akan ada bukti untuk hal ini pada tulang-tulangnya, baik dalam bentuk struktur kaku untuk menopang jumbai tersebut, atau tanda-tanda di tempat di mana otot yang digunakan untuk menggerakkannya menempel. Ia juga menambahkan bahwa apabila hewan ini memang memiliki jumbai, ia tidak akan menggunakannya untuk mengintimidasi makanannya, melainkan untuk mengintimidasi pesaing (ia berspekulasi bahwa hewan itu mungkin bereaksi terhadap karakter dalam film yang menarik tudung ke atas kepalanya).[92] Dalam ulasan tahun 1997 tentang buku mengenai sains di Jurassic Park, ahli paleontologi Peter Dodson juga menunjukkan kesalahan skala ukuran Dilophosaurus dalam film tersebut, serta ketidakmungkinan dari bisa dan jumbainya.[93] Bakker menyoroti pada tahun 2014 bahwa Dilophosaurus di film tersebut tidak memiliki takik yang menonjol di rahang atasnya, dan menyimpulkan bahwa para pembuat film telah melakukan pekerjaan yang baik dalam menciptakan khimaira yang menakutkan dari berbagai hewan yang berbeda, tetapi memperingatkan bahwa hal itu tidak dapat digunakan untuk mengajarkan tentang hewan aslinya.[94] Brown dan Marsh menyatakan bahwa meskipun ciri-ciri ini fiktif, hal tersebut dibuat agar dapat dipercaya dengan mendasarkannya pada biologi hewan nyata.[15] Welles sendiri "sangat senang" melihat Dilophosaurus di Jurassic Park: ia mencatat adanya ketidakakuratan, tetapi menganggapnya sebagai poin kecil, menikmati filmnya, dan merasa bahagia mendapati dinosaurus tersebut menjadi "aktor yang dikenal secara internasional".[95]
Referensi
sunting- ^ "Dilophosaurus". Lexico UK English Dictionary. Oxford University Press. Diarsipkan dari asli tanggal 2020-02-24.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac Welles, S.P. (1984). "Dilophosaurus wetherilli (Dinosauria, Theropoda), osteology and comparisons". Palaeontographica Abteilung A. 185: 85–180.
- ^ a b c Welles, S.P. (1954). "New Jurassic dinosaur from the Kayenta Formation of Arizona". Bulletin of the Geological Society of America. 65 (6): 591–598. Bibcode:1954GSAB...65..591W. doi:10.1130/0016-7606(1954)65[591:NJDFTK]2.0.CO;2.
- ^ Welles, S.P.; Guralnick, R.P. (1994). "Dilophosaurus discovered". ucmp.berkeley.edu. University of California, Berkeley. Diarsipkan dari versi aslinya tanggal November 8, 2017. Diakses tanggal February 13, 2018.
- ^ a b c d e f Naish, D. (2009). The Great Dinosaur Discoveries. London, UK: A & C Black Publishers Ltd. hlm. 94–95. ISBN 978-1-4081-1906-8.
- ^ a b c d e f g h i j k l m n Marsh, A.D.; Rowe, T.B. (2020). "A comprehensive anatomical and phylogenetic evaluation of Dilophosaurus wetherilli (Dinosauria, Theropoda) with descriptions of new specimens from the Kayenta Formation of northern Arizona". Journal of Paleontology. 94 (S78): 1–103. Bibcode:2020JPal...94S...1M. doi:10.1017/jpa.2020.14. S2CID 220601744.
- ^ a b c d e f g Welles, S.P. (1970). "Dilophosaurus (Reptilia: Saurischia), a new name for a dinosaur". Journal of Paleontology. 44 (5): 989. JSTOR 1302738.
- ^ a b Welles, S.P.; Guralnick, R.P. (1994). "Dilophosaurus Details". ucmp.berkeley.edu. University of California, Berkeley. Diarsipkan dari versi aslinya tanggal August 2, 2017. Diakses tanggal February 13, 2018.
- ^ a b Psihoyos, L.; Knoebber, J. (1994). Hunting Dinosaurs. London, UK: Cassell. hlm. 86–89. ISBN 978-0-679-43124-4.
- ^ a b Glut, D.F. (1997). Dinosaurs: The Encyclopedia. Jefferson: McFarland & Company, Inc. hlm. 347–350. ISBN 978-0-7864-7222-2.
- ^ a b Rauhut, O.W. (2004). "The interrelationships and evolution of basal theropod dinosaurs". Special Papers in Palaeontology. 69: 213. Diarsipkan dari versi aslinya tanggal March 24, 2022. Diakses tanggal January 13, 2018.
- ^ a b c d Gay, R. (2001). New specimens of Dilophosaurus wetherilli (Dinosauria: Theropoda) from the early Jurassic Kayenta Formation of northern Arizona. Western Association of Vertebrate Paleontologists Annual Meeting. Vol. 1. Mesa, Arizona. hlm. 1.
- ^ a b c d Carrano, M.T.; Benson, R.B.J.; Sampson, S.D. (2012). "The phylogeny of Tetanurae (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 10 (2): 211–300. Bibcode:2012JSPal..10..211C. doi:10.1080/14772019.2011.630927. S2CID 85354215.
- ^ a b c d Tykoski, R.S. (2005). Anatomy, ontogeny, and phylogeny of coelophysoid theropods (Thesis). University of Texas. hlm. 1–232. Diarsipkan dari versi aslinya tanggal July 29, 2020. Diakses tanggal February 15, 2018 – via UT Libraries: Electronic Theses and Dissertations.
- ^ a b c d e f g h i Brown, M. A.; Marsh, A. D. (2021). "The real Dilophosaurus would have eaten the Jurassic Park version for breakfast". Scientific American. 324 (1): 46–53. doi:10.1038/scientificamerican0121-46. Diarsipkan dari versi aslinya tanggal January 8, 2021. Diakses tanggal January 11, 2021.
- ^ Pickrell, J. (July 7, 2020). "Jurassic Park got almost everything wrong about this iconic dinosaur". National Geographic. Science. Diarsipkan dari asli tanggal July 7, 2020. Diakses tanggal July 12, 2020.
- ^ a b c Gay, R. (2005). "Evidence for sexual dimorphism in the Early Jurassic theropod dinosaur, Dilophosaurus and a comparison with other related forms". Dalam Carpenter, K. (ed.). The Carnivorous Dinosaurs. Bloomington, IN: Indiana University Press. hlm. 277–283. ISBN 978-0-253-34539-4.
- ^ Mortimer, M. (May 2010). "Pickering's taxa 6: Dilophosaurus breedorum". Theropoddatabase.blogspot.com. Diarsipkan dari versi aslinya tanggal December 29, 2017. Diakses tanggal December 29, 2017.
- ^ Hendrickx, C.; Mateus, O.; Evans, Alistair Robert (2014). "Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in non-avian theropods". PLOS ONE. 9 (3) e88905. Bibcode:2014PLoSO...988905H. doi:10.1371/journal.pone.0088905. PMC 3943790. PMID 24598585.
- ^ Hu, S. (1993). "A short report on the occurrence of Dilophosaurus from Jinning County, Yunnan Province". Vertebrata PalAsiatica. 1 (dalam bahasa Tionghoa). 31 (Edisi 1): 65–69.
- ^ a b Lamanna, M.C.; Holtz, T.R. Jr.; Dodson, P. (1998). "A reassessment of the Chinese theropod dinosaur Dilophosaurus sinensis". Journal of Vertebrate Paleontology. Abstracts of Papers, Fifty-Eighth Annual Meeting, Society of Vertebrate Paleontology. 18 (3): 57–58. JSTOR 4523942.
- ^ Xing, L.; Bell, P.R.; Rothschild, B.M.; Ran, H.; Zhang, J.; Dong, Z.; Zhang, W.; Currie, P.J. (2013). "Tooth loss and alveolar remodeling in Sinosaurus triassicus (Dinosauria: Theropoda) from the Lower Jurassic strata of the Lufeng Basin, China". Chinese Science Bulletin. 58 (16): 1931. Bibcode:2013ChSBu..58.1931X. doi:10.1007/s11434-013-5765-7.
- ^ Wang, Guo-Fu; You, Hai-Lu; Pan, Shi-Gang; Wang, Tao (2017). "A new crested theropod dinosaur from the Early Jurassic of Yunnan Province, China". Vertebrata PalAsiatica. 55 (2): 177–186. Diarsipkan dari versi aslinya tanggal August 19, 2021. Diakses tanggal December 31, 2017.
- ^ a b c d e f g h Paul, G.S. (1988). Predatory Dinosaurs of the World. New York, NY: Simon & Schuster. hlm. 258, 267–271. ISBN 978-0-671-61946-6.
- ^ Holtz, T.R. Jr (2012). Dinosaurs: The Most Complete, Up-to-date Encyclopedia for Dinosaur Lovers of All Ages. New York, NY: Random House. hlm. 81. ISBN 978-0-375-82419-7.
- ^ Paul, G.S. (2010). The Princeton Field Guide to Dinosaurs. Princeton, NJ: Princeton University Press. hlm. 75. ISBN 978-0-691-13720-9.
- ^ a b Gierliński, G. (1996). "Feather-like impressions in a theropod resting trace from the Lower Jurassic of Massachusetts". Museum of Northern Arizona Bulletin. 60: 179–184.
- ^ a b Kundrát, M. (2004). "When did theropods become feathered? – Evidence for pre-Archaeopteryx feathery appendages". Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 302B (4): 355–364. Bibcode:2004JEZB..302..355K. doi:10.1002/jez.b.20014. PMID 15287100.
- ^ a b Lockley, M.; Matsukawa, M.; Jianjun, L. (2003). "Crouching theropods in taxonomic jungles: Ichnological and ichnotaxonomic investigations of footprints with metatarsal and ischial impressions" (PDF). Ichnos. 10 (2–4): 169–177. Bibcode:2003Ichno..10..169L. doi:10.1080/10420940390256249. S2CID 128759174.
- ^ a b Martin, A.J.; Rainforth, E.C. (2004). "A theropod resting trace that is also a locomotion trace: case study of Hitchcock's specimen AC 1/7". Geological Society of America. Abstracts with Programs. 36 (2): 96. Diarsipkan dari asli tanggal May 31, 2004.
- ^ a b c d e f g Tykoski, R.S.; Rowe, T. (2004). "Ceratosauria". Dalam Weishampel, D.B.; Dodson, P.; Osmolska, H. (ed.). The Dinosauria (Edisi 2). Berkeley, CA: University of California Press. hlm. 47–70. ISBN 978-0-520-24209-8.
- ^ Welles, S.P. (1983). "Two centers of ossification in a theropod astragalus". Journal of Paleontology. 57 (2): 401. JSTOR 1304663.
- ^ Welles, S.P.; Long, R.A. (1974). "The tarsus of theropod dinosaurs" [Annals of the South African Museum]. Annale van die Suid-Afrikaanse Museum. 64: 191–218. ISSN 0303-2515. Diarsipkan dari versi aslinya tanggal February 8, 2022. Diakses tanggal February 6, 2018.
- ^ Holtz, T.R. Jr. (1994). "The phylogenetic position of the Tyrannosauridae: Implications for theropod systematics". Journal of Paleontology. 68 (5): 1100–1117. Bibcode:1994JPal...68.1100H. doi:10.1017/S0022336000026706. JSTOR 1306180. S2CID 129684676.
- ^ Madsen, J.H.; Welles, S.P. (2000). "Ceratosaurus (Dinosauria, Theropoda): A revised osteology". Utah Geological Survey: 1–89. 41293. Diarsipkan dari asli tanggal January 13, 2018. Diakses tanggal January 13, 2018.
- ^ a b c Yates, A.M. (2005). "A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods". Palaeontologia Africana. 41: 105–122. ISSN 0078-8554. Diarsipkan dari versi aslinya tanggal February 8, 2022. Diakses tanggal February 7, 2018.
- ^ Smith, N.D.; Makovicky, P.J.; Hammer, W.R.; Currie, P.J. (2007). "Osteology of Cryolophosaurus ellioti (Dinosauria: Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution". Zoological Journal of the Linnean Society. 151 (2): 377–421. doi:10.1111/j.1096-3642.2007.00325.x.
- ^ Hendrickx, C.; Hartman, S.A.; Mateus, O. (2015). "An overview of non-avian theropod discoveries and classification". PalArch's Journal of Vertebrate Palaeontology. 12 (1): 73. Diarsipkan dari asli tanggal June 22, 2018. Diakses tanggal January 13, 2018.
- ^ Zahner, M.; Brinkmann, W. (2019). "A Triassic averostran-line theropod from Switzerland and the early evolution of dinosaurs". Nature Ecology & Evolution. 3 (8): 1146–1152. Bibcode:2019NatEE...3.1146Z. doi:10.1038/s41559-019-0941-z. PMC 6669044. PMID 31285577.
- ^ a b Welles, S.P. (1971). "Dinosaur footprints from the Kayenta Formation of northern Arizona". Plateau. 44: 27–38.
- ^ Morales, M.; Bulkley, S. (1996). "Paleoichnological evidence for a theropod dinosaur larger than Dilophosaurus in the Lower Jurassic Kayenta Formation. The Continental Jurassic". Museum of Northern Arizona Bulletin. 60: 143–145.
- ^ Gierliński, G. (1991). "New dinosaur ichnotaxa from the Early Jurassic of the Holy Cross Mountains, Poland". Palaeogeography, Palaeoclimatology, Palaeoecology. 85 (1–2): 137–148. Bibcode:1991PPP....85..137G. doi:10.1016/0031-0182(91)90030-U.
- ^ Gierliński, G.; Ahlberg, A. (1994). "Late triassic and early jurassic dinosaur footprints in the Höganäs Formation of southern Sweden". Ichnos. 3 (2): 99. Bibcode:1994Ichno...3...99G. doi:10.1080/10420949409386377.
- ^ Gierliński, G.; Sabath, K. (2005). "The Hitchcock enigma: first feathers or artifacts" (PDF). Tracking Dinosaur Origins: The Triassic/Jurassic Terrestrial Transition. hlm. 21–22.
- ^ Weems, R.E. (2003). "Plateosaurus foot structure suggests a single trackmaker for Eubrontes and Gigandipus footprints". Dalam le Tourneau, P.M.; Olsen, P.E. (ed.). The Great Rift Valleys of Pangea in Eastern North America. Vol. 2. New York: Columbia University Press. hlm. 293–313. ISBN 978-0-231-12676-2.
- ^ Rainforth, E.C. (2003). "Revision and re-evaluation of the Early Jurassic dinosaurian ichnogenus Otozoum". Palaeontology. 46 (4): 803–838. Bibcode:2003Palgy..46..803R. doi:10.1111/1475-4983.00320.
- ^ a b Weems, R.E. (2006). "The manus print of Kayentapus minor; its bearing on the biomechanics and ichnotaxonomy of Early Mesozoic saurischian dinosaurs". New Mexico Museum of Natural History and Science Bulletin. 37: 369–378.
- ^ Lucas, S.G.; Klein, H.; Lockley, M.G.; Spielmann, J.A.; Gierlinski, G.D.; Hunt, A. P.; Tanner, L.H. (2006). "Triassic–Jurassic stratigraphic distribution of the theropod footprint ichnogenus Eubrontes". New Mexico Museum of Natural History and Science Bulletin. 37. 265. Diarsipkan dari versi aslinya tanggal October 4, 2016. Diakses tanggal February 7, 2019.
- ^ a b c Milner, Andrew R.C.; Harris, J.D.; Lockley, M.G.; Kirkland, J.I.; Matthews, N.A.; Harpending, H. (2009). "Bird-like anatomy, posture, and behavior revealed by an Early Jurassic theropod dinosaur resting trace". PLOS ONE. 4 (3) e4591. Bibcode:2009PLoSO...4.4591M. doi:10.1371/journal.pone.0004591. PMC 2645690. PMID 19259260.
- ^ Weems, R.E. (2019). "Evidence for bipedal prosauropods as the likely Eubrontes track-makers". Ichnos. 26 (3): 187–215. Bibcode:2019Ichno..26..187W. doi:10.1080/10420940.2018.1532902. S2CID 133770251.
- ^ Lucas, Spencer G. (2024). "Jurassic vertebrate tracks and traces". Vertebrate Ichnology: Tetrapod Tracks and Trackways (Edisi 1st). Chantilly: Elsevier. hlm. 331. ISBN 978-0-443-13837-9.
- ^ a b Bakker, R.T. (1986). The Dinosaur Heresies. New York, NY: William Morrow. hlm. 263–264, 339. ISBN 978-0-8217-5608-9.
- ^ Therrien, F.; Henderson, D.; Ruff, C. (2005). "Bite me – biomechanical models of theropod mandibles and implications for feeding behavior". Dalam Carpenter, K. (ed.). The Carnivorous Dinosaurs. Indiana University Press. hlm. 179–230. ISBN 978-0-253-34539-4.
- ^ Milner, A.; Kirkland, J. (2007). "The case for fishing dinosaurs at the St. George Dinosaur Discovery Site at Johnson Farm" (PDF). Survey Notes of the Utah Geological Survey. 39: 1–3. Diarsipkan dari asli (PDF) tanggal November 4, 2017. Diakses tanggal May 14, 2015.
- ^ Marsh, A.D.; Rowe, T.B. (2018). "Anatomy and systematics of the sauropodomorph Sarahsaurus aurifontanalis from the Early Jurassic Kayenta Formation". PLOS ONE. 13 (10) e0204007. Bibcode:2018PLoSO..1304007M. doi:10.1371/journal.pone.0204007. PMC 6179219. PMID 30304035.
- ^ a b Czerkas, S.J.; Czerkas, S.A. (1990). Dinosaurs: A global view. Limpsfield: Dragons' World. hlm. 208. ISBN 978-0-7924-5606-3.
- ^ Christiansen, Per (1998). "Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential". Gaia. 15: 241–255. ISSN 0871-5424. S2CID 83115549.
- ^ Paul, G.S., ed. (2000). "Color section". The Scientific American Book of Dinosaurs. New York, NY: St. Martin's Press. hlm. 216. ISBN 978-0-312-31008-0.
- ^ Senter, P.; Robins, J.H. (2005). "Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis, and implications for predatory behaviour". Journal of Zoology. 266 (3): 307–318. doi:10.1017/S0952836905006989.
- ^ Senter, P.; Robins, J.H. (2015). "Resting orientations of dinosaur scapulae and forelimbs: A numerical analysis, with implications for reconstructions and museum mounts". PLOS ONE. 10 (12) e0144036. Bibcode:2015PLoSO..1044036S. doi:10.1371/journal.pone.0144036. PMC 4684415. PMID 26675035.
- ^ a b c d Senter, P.; Sullivan, C. (2019). "Forelimbs of the theropod dinosaur Dilophosaurus wetherilli: Range of motion, influence of paleopathology and soft tissues, and description of a distal carpal bone". Palaeontologia Electronica. doi:10.26879/900.
- ^ Coombs, W.P. (1990). "Behavior patterns of dinosaurs". Dalam Weishampel, D.B.; Osmolska, H.; Dodson, P. (ed.). The Dinosauria (Edisi 1st). Berkeley, CA: University of California Press. hlm. 42. ISBN 978-0-520-06727-1.
- ^ Padian, K.; Horner, J.R. (2011). "The evolution of 'bizarre structures' in dinosaurs: Biomechanics, sexual selection, social selection or species recognition?". Journal of Zoology. 283 (1): 3–17. doi:10.1111/j.1469-7998.2010.00719.x.
- ^ Knell, R.J.; Sampson, S. (2011). "Bizarre structures in dinosaurs: species recognition or sexual selection? A response to Padian and Horner" (PDF). Journal of Zoology. 283 (1): 18–22. doi:10.1111/j.1469-7998.2010.00758.x. Diarsipkan (PDF) dari versi aslinya tanggal July 22, 2018. Diakses tanggal August 6, 2018.
- ^ Hone, D.W.E.; Naish, D. (2013). "The 'species recognition hypothesis' does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs". Journal of Zoology. 290 (3): 172–180. doi:10.1111/jzo.12035.
- ^ a b Tkach, J.S. (October 19, 1996). Multi-element osteohistological analysis of Dilphosaurus wetherilli. Fifty-sixth Annual Meeting, Society of Vertebrate Paleontology. Journal of Vertebrate Paleontology. Abstracts of Papers. Vol. 16, no. 3. American Museum of Natural History, New York, New York. hlm. 1A – 80A. JSTOR 4523755.
- ^ a b Molnar, R.E. (2001). "Theropod paleopathology: A literature survey". Dalam Tanke, D.H.; Carpenter, K. (ed.). Mesozoic Vertebrate Life. Bloomington, IN: Indiana University Press. hlm. 337–363. ISBN 978-0-253-33907-2.
- ^ a b Rothschild, B.; Tanke, D.H.; Ford, T.L. (2001). "Theropod stress fractures and tendon avulsions as a clue to activity". Dalam Tanke, D.H.; Carpenter, K. (ed.). Mesozoic Vertebrate Life. Bloomington, IN: Indiana University Press. hlm. 331–336. ISBN 978-0-253-33907-2.
- ^ a b c d e f Senter, P.; Juengst, S.L.; Heymann, D. (2016). "Record-breaking pain: The largest number and variety of forelimb bone maladies in a theropod dinosaur". PLOS ONE. 11 (2) e0149140. Bibcode:2016PLoSO..1149140S. doi:10.1371/journal.pone.0149140. PMC 4765892. PMID 26909701.
- ^ Marsh, Adam Douglas (2018). Contextualizing the evolution of theropod dinosaurs in western North America using U-Pb geochronology of the Chinle Formation and Kayenta Formation on the Colorado Plateau (Thesis). doi:10.26153/tsw/41876 (tidak aktif July 12, 2025). Pemeliharaan CS1: DOI nonaktif per Juli 2025 (link)
- ^ Steiner, Maureen; Tanner, Lawrence H. (2014). "Magnetostratigraphy and paleopoles of the Kayenta Formation and the Tenney Canyon Tongue". Volumina Jurassica. 12 (2): 31–38. doi:10.5604/17313708.1130125 (tidak aktif July 12, 2025). ISSN 1731-3708. Pemeliharaan CS1: DOI nonaktif per Juli 2025 (link)
- ^ a b Harshbarger, J. W.; Repenning, C. A.; Irwin, J. H. (1957). "Stratigraphy of the uppermost Triassic and the Jurassic rocks of the Navajo country". New Mexico Geological Society. 291: 98–114. Diarsipkan dari asli tanggal February 16, 2020. Diakses tanggal February 14, 2018.
- ^ a b c Lucas, S.G.; Heckert, A. B.; Tanner, L.H. (2005). "Arizona's Jurassic fossil vertebrates and the age of the Glen Canyon Group". New Mexico Museum of Natural History Bulletins. 29: 95–104. Diarsipkan dari versi aslinya tanggal November 18, 2016. Diakses tanggal February 15, 2018.
- ^ a b c Luttrell, P.R.; Morales, M. (1993). "Bridging the gap across Moenkopi Wash: a lithostratigraphic correlation". Museum of Northern Arizona Bulletin. 59: 111–127.
- ^ a b Jenkins, F.A.; Shubin, N. H. (1998). "Prosalirus bitis and the anuran caudopelvic mechanism". Journal of Vertebrate Paleontology. 18 (3): 495–510. Bibcode:1998JVPal..18..495J. doi:10.1080/02724634.1998.10011077.
- ^ Rigby, J.K.; Hamblin, W.K.; Matheny, R.; Welsh, S.L. (1971). "Guidebook to the Colorado river: Part 3, Moab to Hite, Utah through Canyonlands National Park" (PDF). Brigham Young University Research Studies, Geology Series. 18 (2): 7. Diarsipkan dari asli (PDF) tanggal November 25, 2020. Diakses tanggal February 15, 2018.
- ^ Lucas, S.G.; Tanner, L.H. (2007). "Tetrapod biostratigraphy and biochronology of the Triassic–Jurassic transition on the southern Colorado Plateau, USA". Palaeogeography, Palaeoclimatology, Palaeoecology. 244 (1–4): 242–256. Bibcode:2007PPP...244..242L. doi:10.1016/j.palaeo.2006.06.030.
- ^ Curtis, K.; Padian, K. (1999). "An Early Jurassic microvertebrate fauna from the Kayenta Formation of northeastern Arizona: microfaunal change across the Triassic-Jurassic boundary". PaleoBios. 19 (2): 19–37.
- ^ a b Jenkins, F. A.; Crompton, A. W.; Downs, W. R. (1983). "Mesozoic mammals from Arizona: New evidence on mammalian evolution". Science. 222 (4629): 1233–1235. Bibcode:1983Sci...222.1233J. doi:10.1126/science.222.4629.1233. PMID 17806725. S2CID 35607107.
- ^ Gay, R. (2010). "Kayentavenator elysiae, a new tetanuran from the early Jurassic of Arizona". Dalam Gay, R. (ed.). Notes on Early Mesozoic Theropods. Lulu Press. hlm. 27–43. ISBN 978-0-557-46616-0.
- ^ Rowe, T.B.; Sues, H.-D.; Reisz, R.R. (2010). "Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon". Proceedings of the Royal Society B: Biological Sciences. 278 (1708): 1044–1053. doi:10.1098/rspb.2010.1867. PMC 3049036. PMID 20926438.
- ^ Hamblin, A.H.; Foster, J.R. (2000). "Ancient animal footprints and traces in the Grand Staircase–Escalante National Monument, south-central Utah". Dalam Sprinkel, D.A.; Chidsey, T.C. Jr.; Anderson, P.B. (ed.). Geology of Utah's Parks and Monuments. Salt Lake City: Utah Geological Association. hlm. 557–568. ISBN 978-1-882054-10-7.
- ^ Reisdorf, A.G.; Wuttke, M. (2012). "Re-evaluating Moodie's opisthotonic-posture hypothesis in fossil vertebrates, Part I: Reptiles – the taphonomy of the bipedal dinosaurs Compsognathus longipes and Juravenator starki from the Solnhofen archipelago (Jurassic, Germany)". Palaeobiodiversity and Palaeoenvironments. 92 (1): 119–168. Bibcode:2012PdPe...92..119R. doi:10.1007/s12549-011-0068-y. S2CID 129785393.
- ^ a b c d Mayor, A. (2005). "The Southwest: Fossil fetishes and monster slayers". Fossil Legends of the First Americans. Princeton, NJ: Princeton University Press. hlm. 130–143. ISBN 978-0-691-13049-1. JSTOR j.ctt4cgcs9.10.
- ^ Stone, M. (2017). "Connecticut welcomes its new state dinosaur". CTBoom.com. Diarsipkan dari asli tanggal September 25, 2017. Diakses tanggal February 13, 2018.
- ^ Rogers, O. (2016). "Discovered dinosaur tracks re-route highway and lead to state park". Connecticut History. Diarsipkan dari versi aslinya tanggal April 11, 2022. Diakses tanggal January 3, 2018.
- ^ Moeser, C. (April 26, 1998). "Arizona dinosaur debate becomes real Godzilla". The Arizona Republic. Deseret News. Diarsipkan dari versi aslinya tanggal May 29, 2022. Diakses tanggal May 12, 2020.
- ^ Gebers, S. (February 19, 2018). "Arizona has no official state dinosaur, so an 11 year-old proposed one – the Sonorasaurus". The Republic. Diakses tanggal February 20, 2018 – via AZ Central.
- ^ Crichton, M. (1990). Jurassic Park. London: Random Century Group. hlm. 76–78. ISBN 978-0-394-58816-2.
- ^ Shay, D.; Duncan, J. (1993). The Making of Jurassic Park. New York: Boxtree Ltd. hlm. 24, 35–36, 113. ISBN 978-1-85283-774-7.
- ^ Duncan, J. (2007). The Winston Effect: The art and history of Stan Winston studio. London, UK: Titan Books. hlm. 177–178. ISBN 978-1-84576-150-9.
- ^ Bennington, J.B. (1996). "Errors in the movie Jurassic Park". American Paleontologist. 4 (2): 4–7. Diarsipkan dari asli tanggal October 2, 2018. Diakses tanggal December 28, 2017.
- ^ Dodson, P. (1997). "Raising the dead". Science. 277 (5326): 644–645. doi:10.1126/science.277.5326.644b. JSTOR 2893254. S2CID 178195275.
- ^ Bakker, R.T. (2014). "A tale of two compys: What Jurassic Park got right – and wrong – about dino anatomy". blog.hmns.org. The Houston Museum of Natural Science. Diarsipkan dari versi aslinya tanggal September 21, 2020. Diakses tanggal August 18, 2020.
- ^ Welles, S.P.; Guralnick, R.P. (1994). "Dilophosaurus, the actor". ucmp.berkeley.edu. University of California, Berkeley. Diarsipkan dari versi aslinya tanggal January 4, 2018. Diakses tanggal February 13, 2018.
Pranala luar
sunting- "Dilophosaurus! A Narrated Exhibition". Bersama Samuel P. Welles. Pemeliharaan CS1: Lain-lain (link)
